生物氧化与氧化磷酸化
生物氧化
呼吸链概述
呼吸链,也叫电子传递链,指一系列蛋白质复合物和其他电子载体,将电子从电子供体传递到电子受体,并将这种氧化还原反应与跨膜质子转运耦联起来。
整个氧化磷酸化过程除了电子传递链以外,还有ATP合成酶,负责释放电子传递链建立的质子梯度驱动ATP合成。
排除歧义
中文语境下,呼吸链特指线粒体的电子传递链,而电子传递链概念更广泛包括原核与光合作用等的电子传递过程。
线粒体超微结构
- 线粒体是双层膜细胞器,外膜存在孔蛋白(Porin)可以通透10kDa以下的分子,而内膜具有选择通透性,载体蛋白存在于内膜。
- 内膜向内折叠形成嵴(Cristae),呼吸链复合物和ATP合成酶存在于嵴上。
- 膜间隙(Intermembrane space, IMS)为正电位,基质(Matrix)为负电位,质子向IMS转运相当于建立质子梯度。
通用电子载体
通用电子载体可以作为氧化剂或者还原剂参与许多氧化还原的代谢反应,主要有两种:
- 辅酶:可以游离于酶存在,在不同底物/酶之间穿梭传递电子:NAD+, NADP+, CoQ
- 辅基:紧密甚至共价结合在酶内部,仅能为该酶催化的反应传递电子:FMN, FAD, 铁硫簇, 血红素, 铜离子.
- 小型可溶蛋白质:内部结合有充当电子载体的辅基:细胞色素c(内含血红素),铁氧还蛋白(内含铁硫簇),质体蓝素(内含铜离子)
根据携带电子数量的不同,分为:
- 双电子载体: 内部结合有充当电子载体的辅基:细胞色素c(内含血红素),铁氧还蛋白(内含铁硫簇),质体蓝素(内含铜离子)
- 单或双电子载体:黄素单核苷酸(FMN)和黄素腺嘌呤二核苷酸(FAD)
NAD / NADP
呼吸链中的其他电子载体
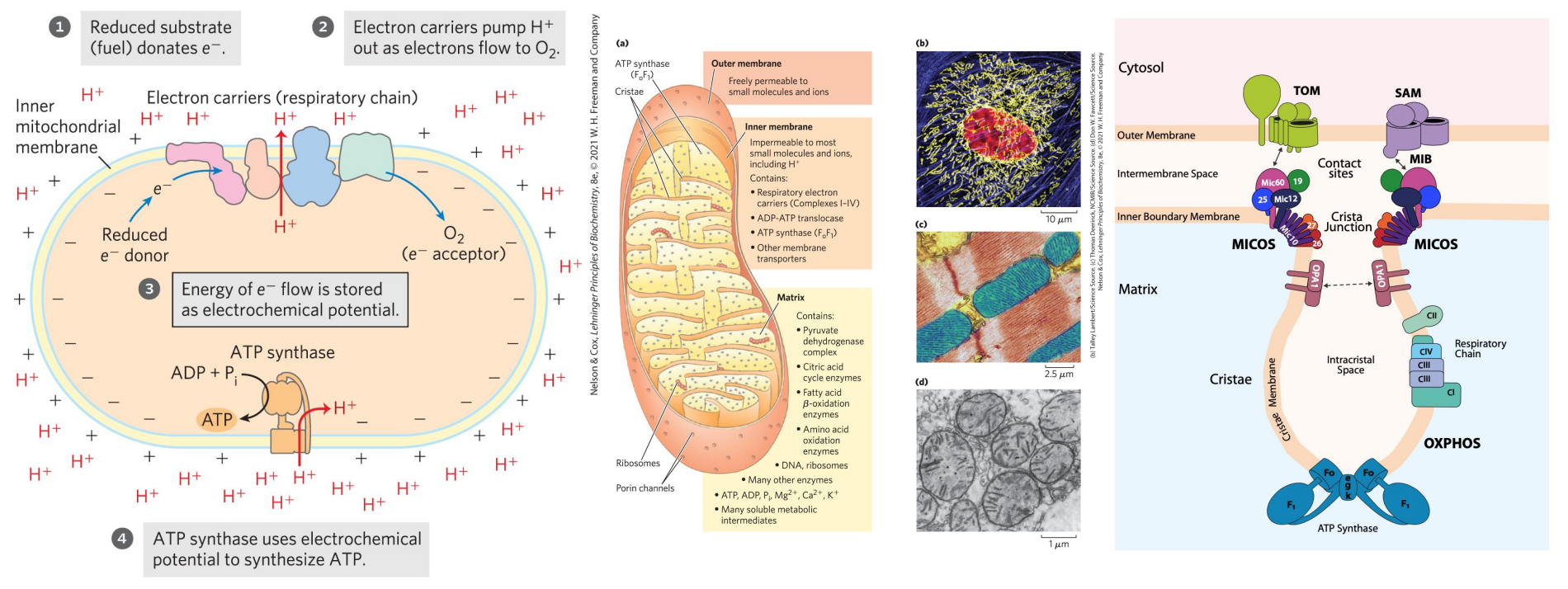
泛醌/辅酶Q(Co-enzyme Q10,Ubiquinone, CoQ,CoQ10,UQ, Q10, Q),其具有一个苯醌头部与10聚戊二烯尾部组成,脂溶性辅酶,可在线粒体内膜中自由扩散 ,能进行可逆的氧化还原反应,呼吸链中唯一不与蛋白质结合的电子载体。
其负责传递电子和氢,处在电子传递的中心地位,接受一个电子和一个质子还原成半醌,再接受一个电子和一个质子还原成二氢泛醌,后者又可脱去电子和质子而被氧化为泛醌。
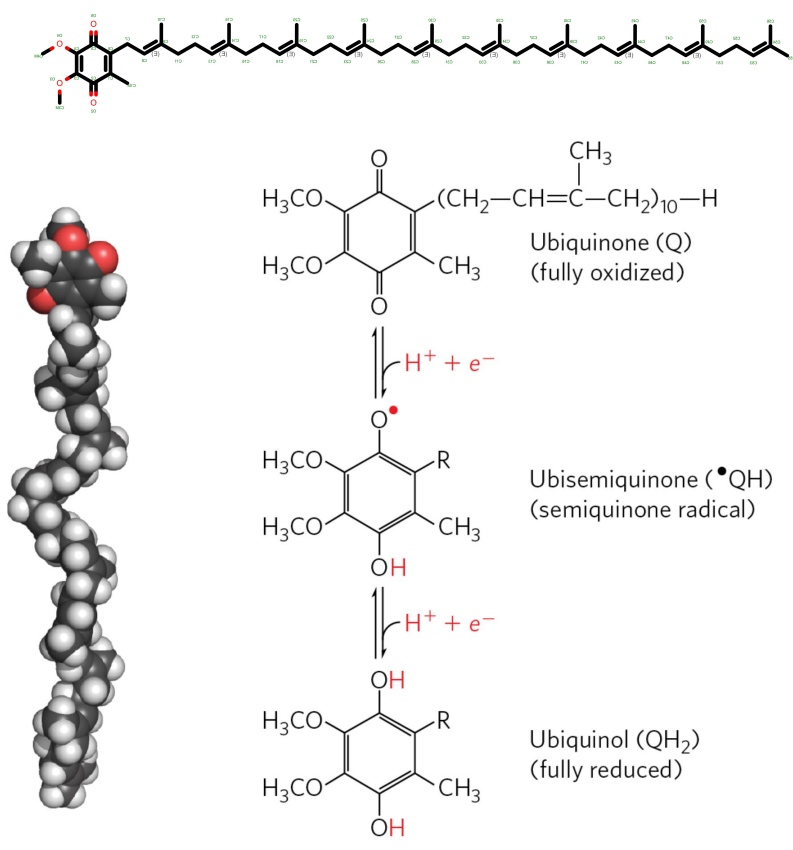
铁硫蛋白中辅基铁硫中心(Fe-S)含有等量铁原子和硫原子,其中一个铁原子可进行\(\ce{Fe^{2+} \rightleftharpoons Fe^{3+} + e-}\)反应传递电子,属于单电子传递体。
氧化还原电位
高中我们学过:
这个口诀描述了不同金属单质之间还原性的强弱差异,本质上是氧化还原电位的差异。这与我们在普化中学习到的电化学中的原理有关(与标准氢电极相比)。
一般电子只能自发从低电位转移至高电位,过程中发生的氧化还原反应释放的能量为\(\Delta G = -nF\Delta E_h\)(n为专业电子数字,F为法拉第常数)。
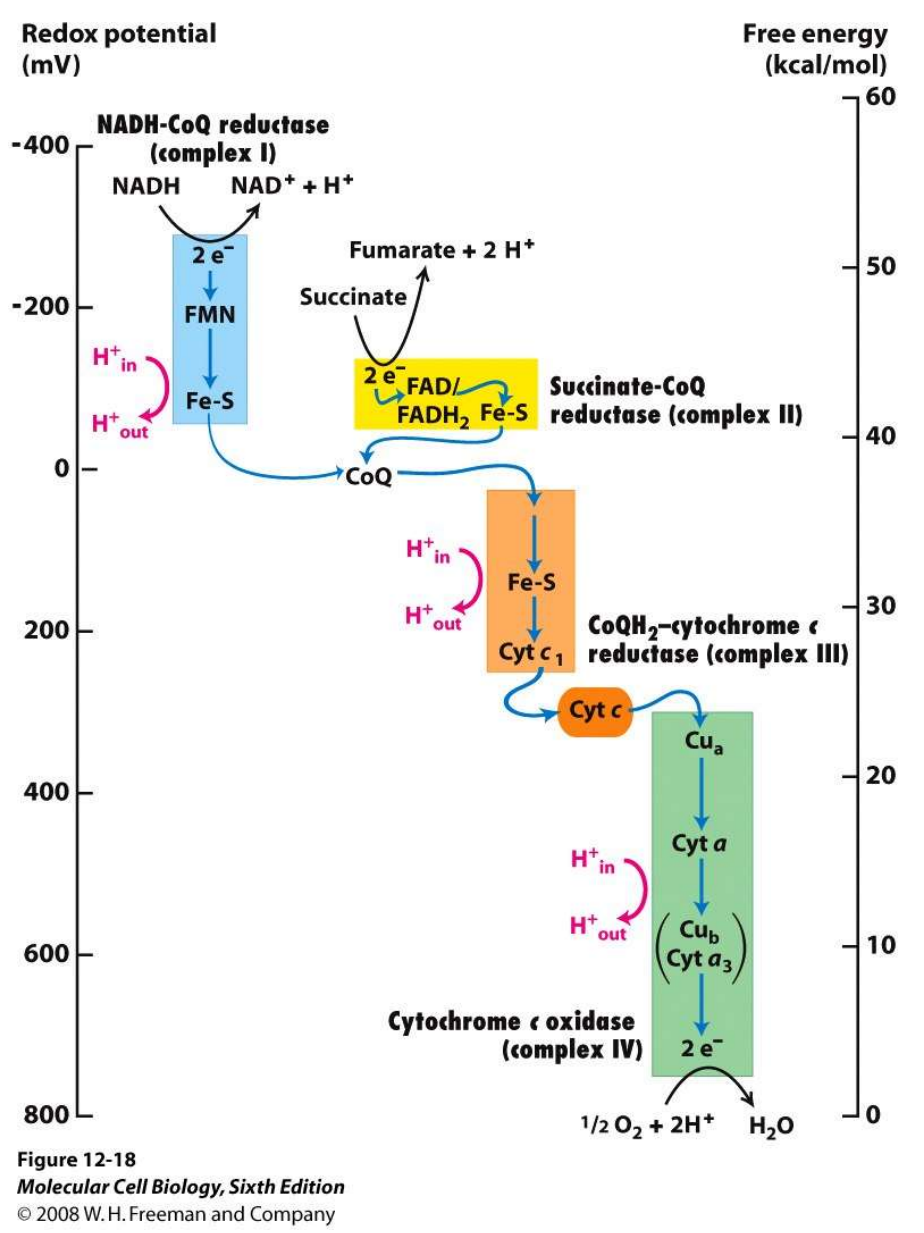
呼吸链的电子传递过程如下:
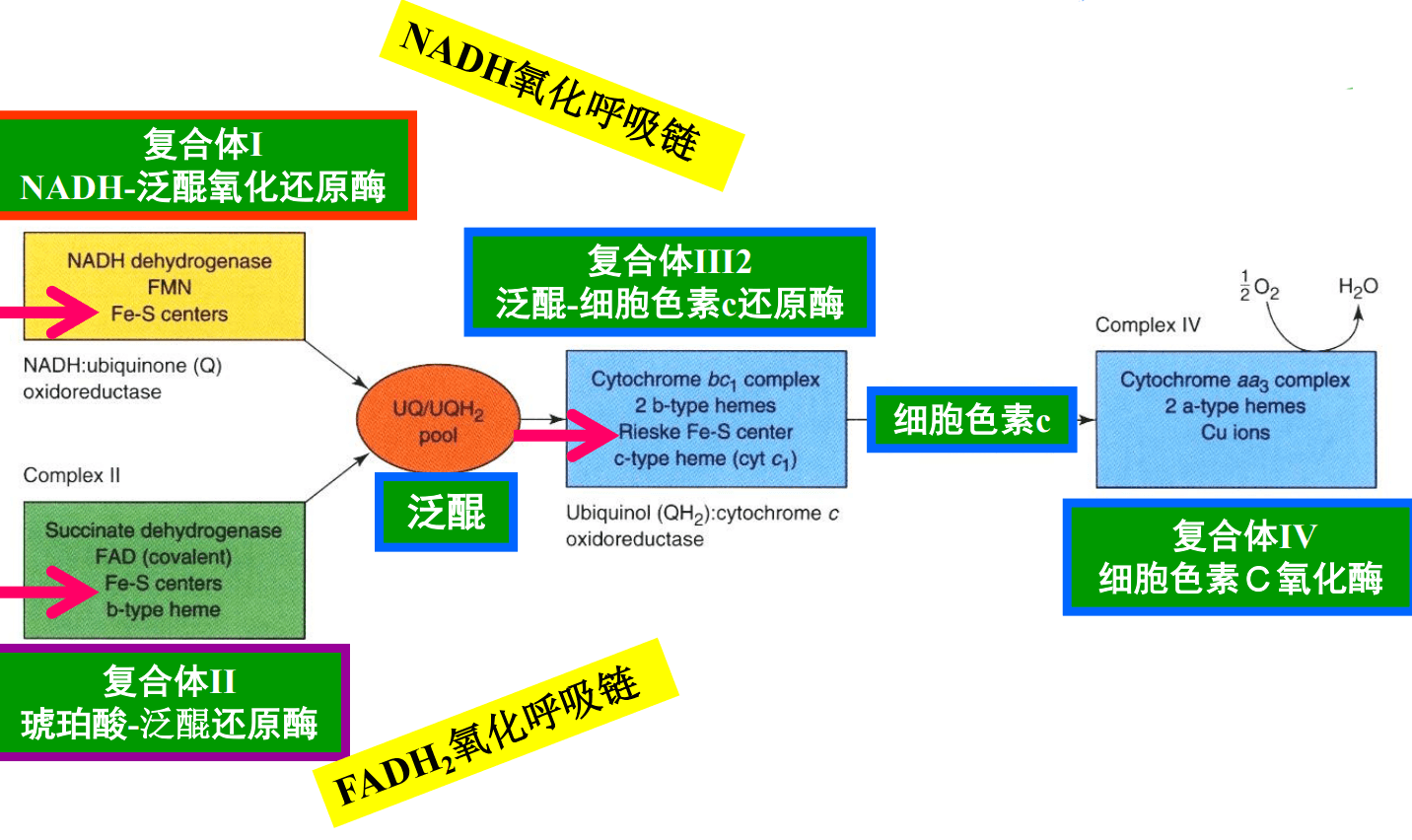
复合物III起到了承下启下的中枢作用,将来自复合体I的\(\ce{NADH}\)和复合体II的\(\ce{FADH2}\)的电子汇总,传递给复合体IV,最终传递给氧气。
呼吸链复合物
<待补充>
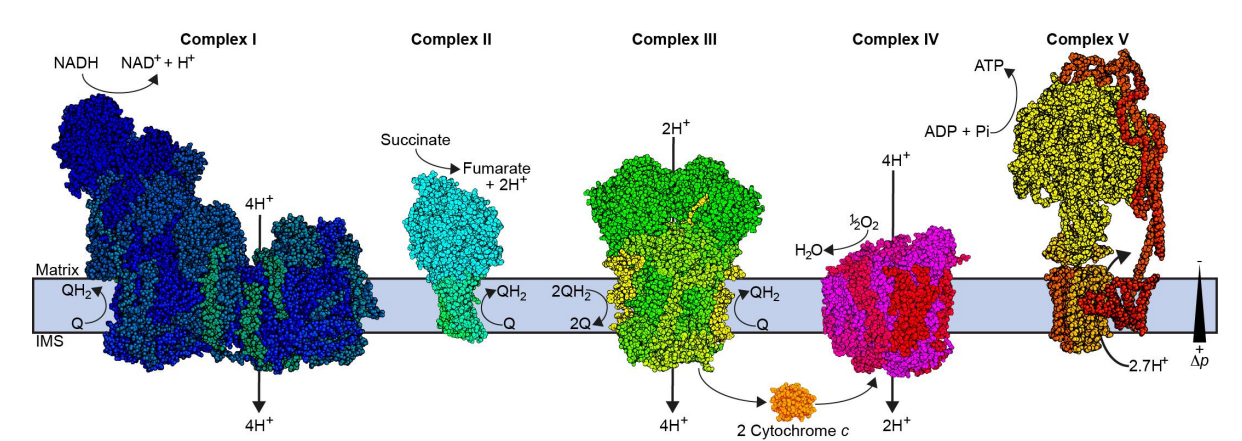
复合物I
复合物I,也称NADH脱氢酶、NADH:CoQ氧还酶、NDH-1,是哺乳动物电子传递链中首个也是最大的跨膜复合物。
复合物I结构组成如下:
- 两个结构臂:跨膜臂 (Membrane arm),基质臂 (Peripheral/Matrixarm)
- 14个核心亚基:7个在基质臂,7个在跨膜臂(仅跨膜核心亚基由线粒体基因组编码)
- 传电子辅基:FMN,铁硫簇
- 电子供体与受体:NADH,CoQ
值得注意的是原核生物CI仅有核心亚基,真核生物CI还包括数量不等的附属亚基。
膜外电子传递
CI首先进行膜外电子传递,将NADH上拔下来的电子一步步传递给泛醌。
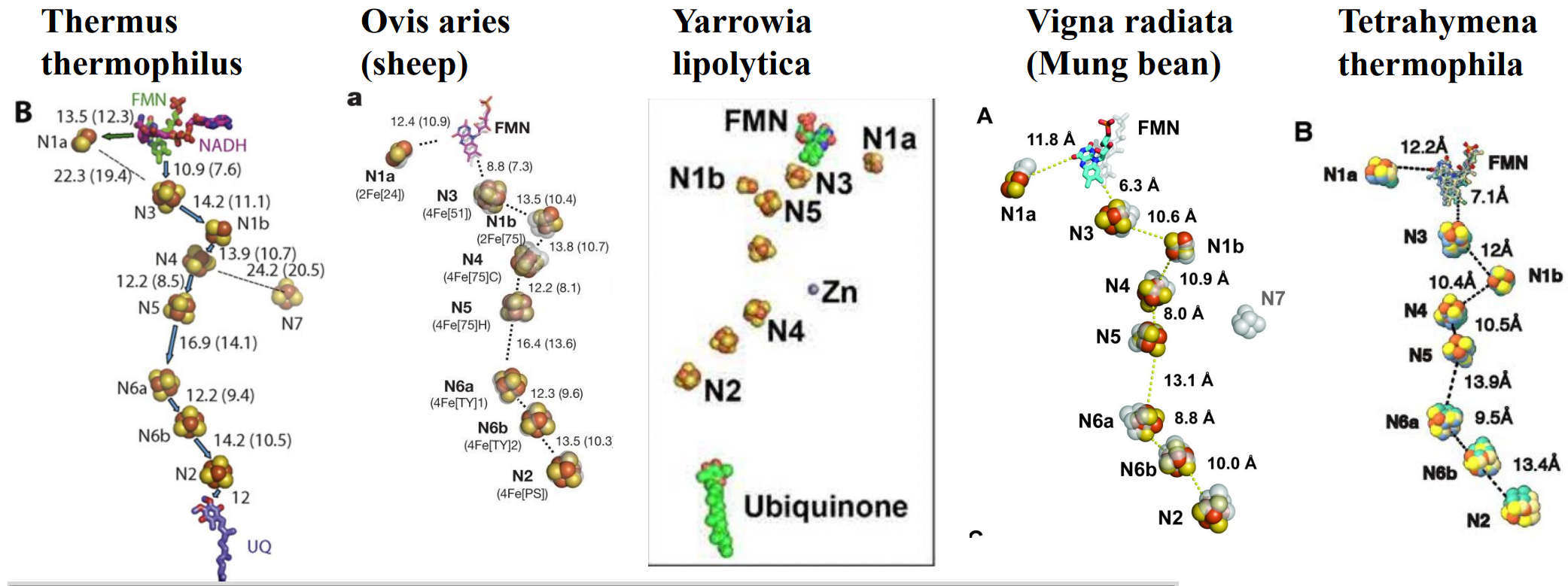
CoQ还原与质子跨膜转运
ND2,ND4,ND5具有Na+-H+ antiporter结构,可以跨膜转运质子。ND1,ND3,ND4L,ND6位于膜内臂和基质臂交界处,其中ND1,ND3和基质臂的NDUFS2,NDUFS7共同组成CoQ进出复合物I的通道,而ND6起到耦联电子传递和质子转运的功能。
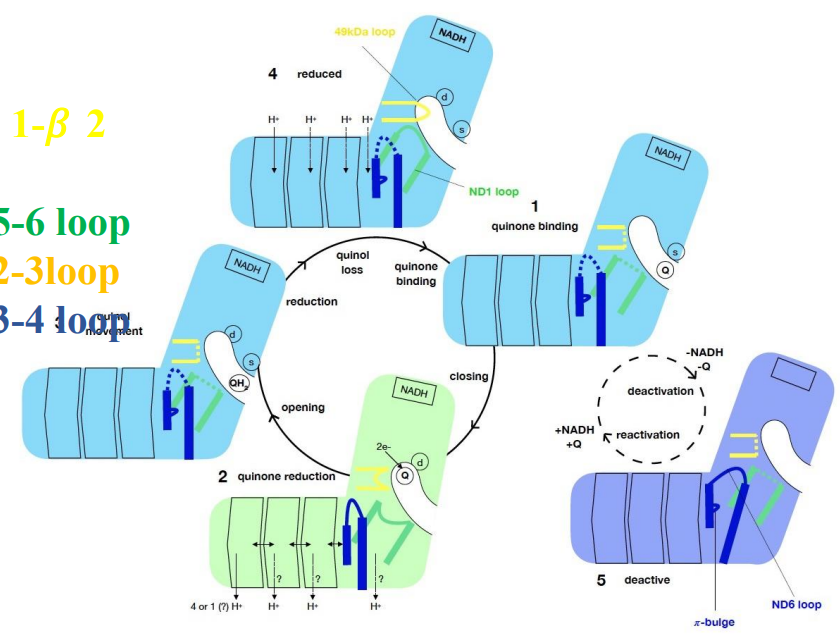
关键的问题是:电子传递与质子跨膜转运如何耦联?
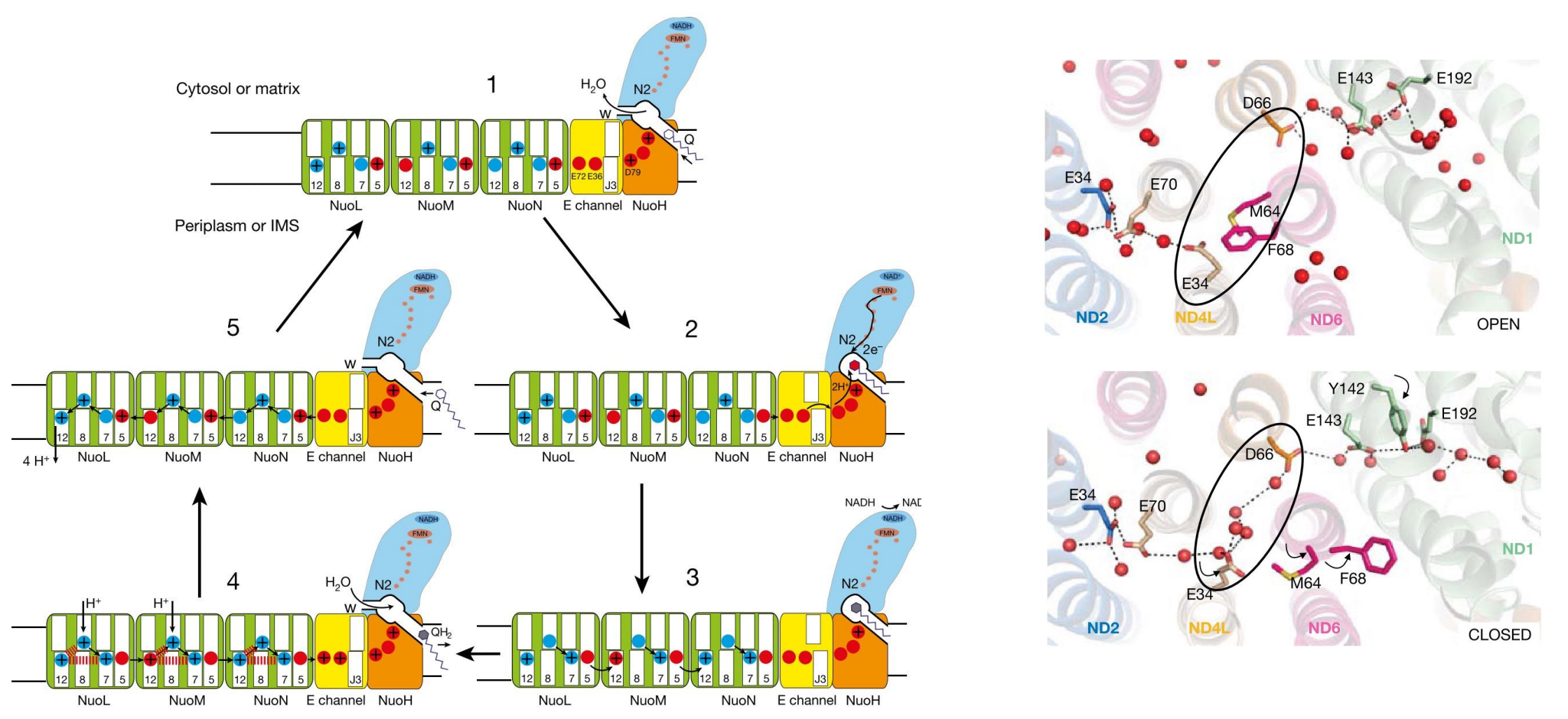
有一个比较被广泛接受的假说,由Sazanov等提出,其认为:
- CI的整个催化周期存在“开放-闭合”的构象转变。
- 此种构象转变使得ND6的TMH3出现“π bulge-full helix”的变化,从而使得跨膜臂亲水轴出现断续的变化。
- CoQ被还原需要从附近氨基酸获得质子,产生的静电波动诱发亲水轴的质子发生重排。
个人理解
感觉就像是一个“打羽毛球”的过程,一开始M64与F68就绪 (CLOSED),D66和E34形成亲水轴,吸引氢离子。
电子传递开始,释放的化学能驱动M64与F68开始转动,像打羽毛球一样“击中”并“切断”了亲水轴,一拍把氢离子拍飞了出去。
复合物II
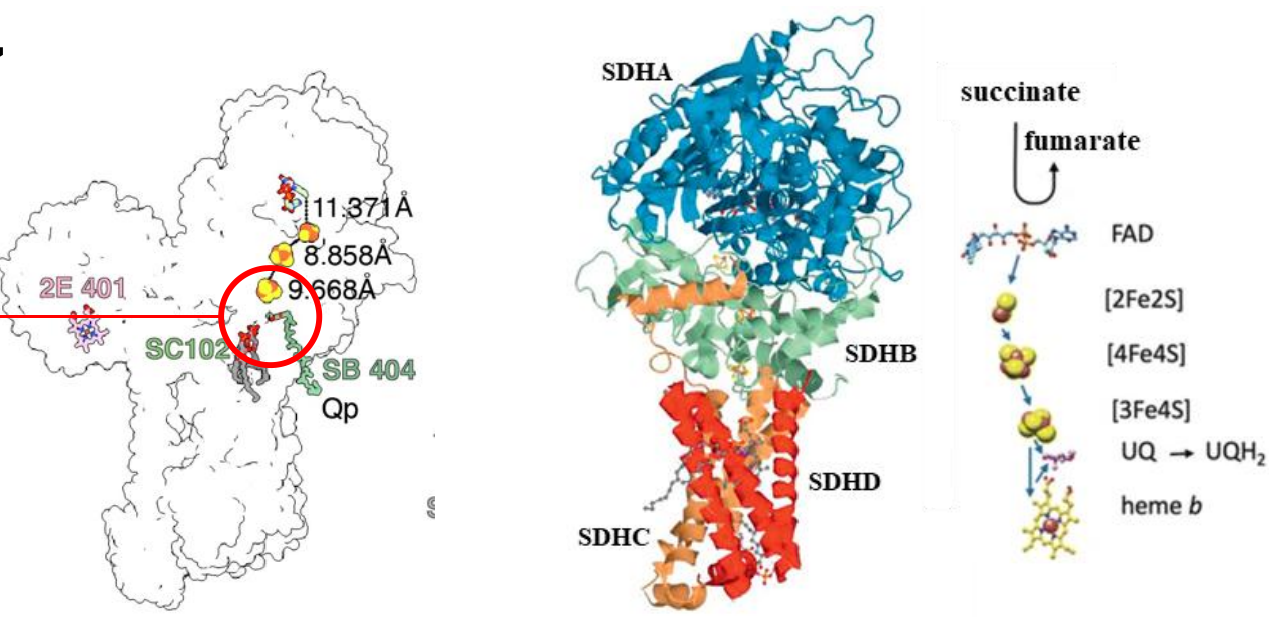
复合物II是电子传递链的另一个入口,从琥珀酸处获得电子。
- 复合物结构包含四个核心亚基:
- 基质侧亲水亚基SDHA,SDHB
- 跨膜疏水亚基SDHC,SDHD
- 传电子辅基:
- 共价结合的FAD
- 依次存在\(\ce{2Fe2S}\)簇,\(\ce{4Fe4S}\)簇和\(\ce{3Fe4S}\)簇,\(\ce{3Fe4S}\)簇直接将电子交给CoQ
- 膜内有时存在heme b,但不参与电子传递
- 不能跨膜转运质子
CoQ多电子途径还原
复合体V
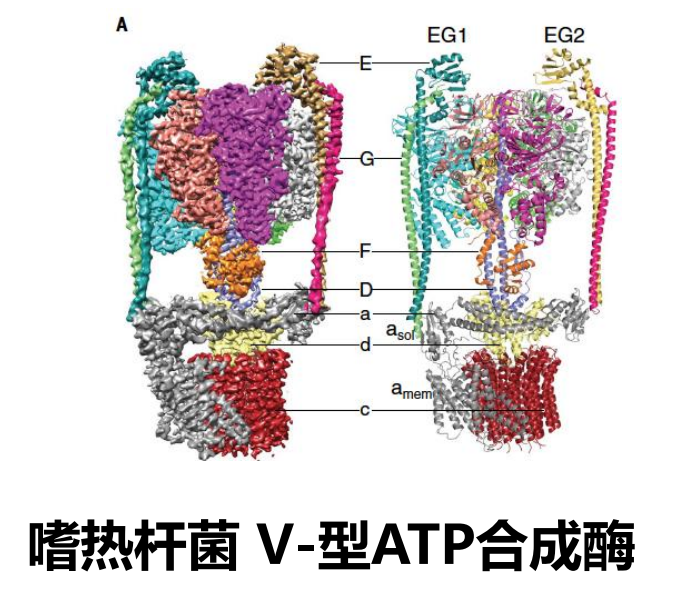
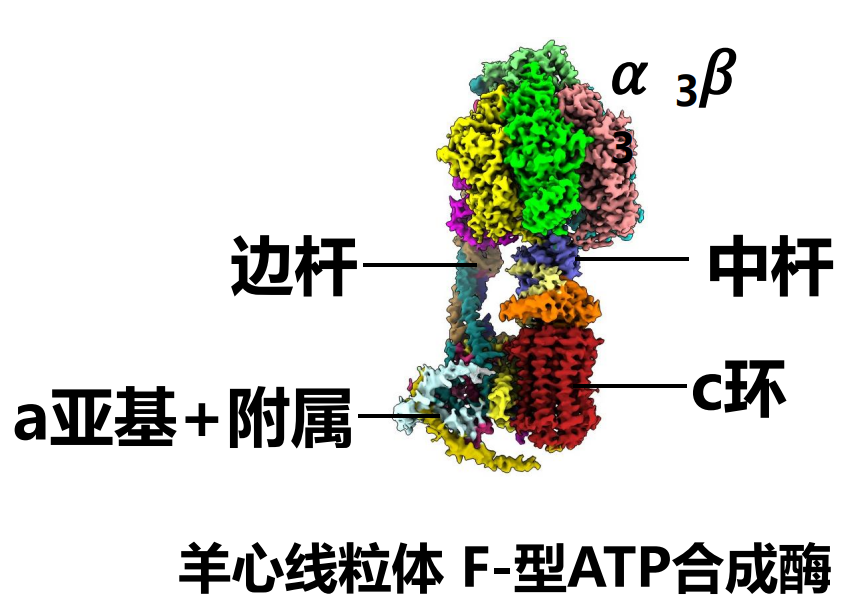
复合体V,也就是F型ATP合成酶,其整体包含两大部分:膜外 (F1) 部分和膜内 (F0) 部分 (也可以分为转子和定子两部分)。
细分来说,又分为\(\alpha_3\beta_3\),中杆 (centralstalk),边杆 (peripheralstalk),c环 (c ring),a亚基 (subunit a) 及其附属亚基。其中F1包含\(\alpha_3\beta_3\),中杆,边杆的一部分(如果是V型则是全部)。F0包含c环,a亚基及其附属亚基。
ATP合成酶中,转子是中杆与c 环,而定子是\(\alpha_3\beta_3\),边杆,a亚基及其附属亚基。
工作原理——旋转催化
ATP合成酶是如何合成ATP的?它是“转着”合成的!(大风车?)
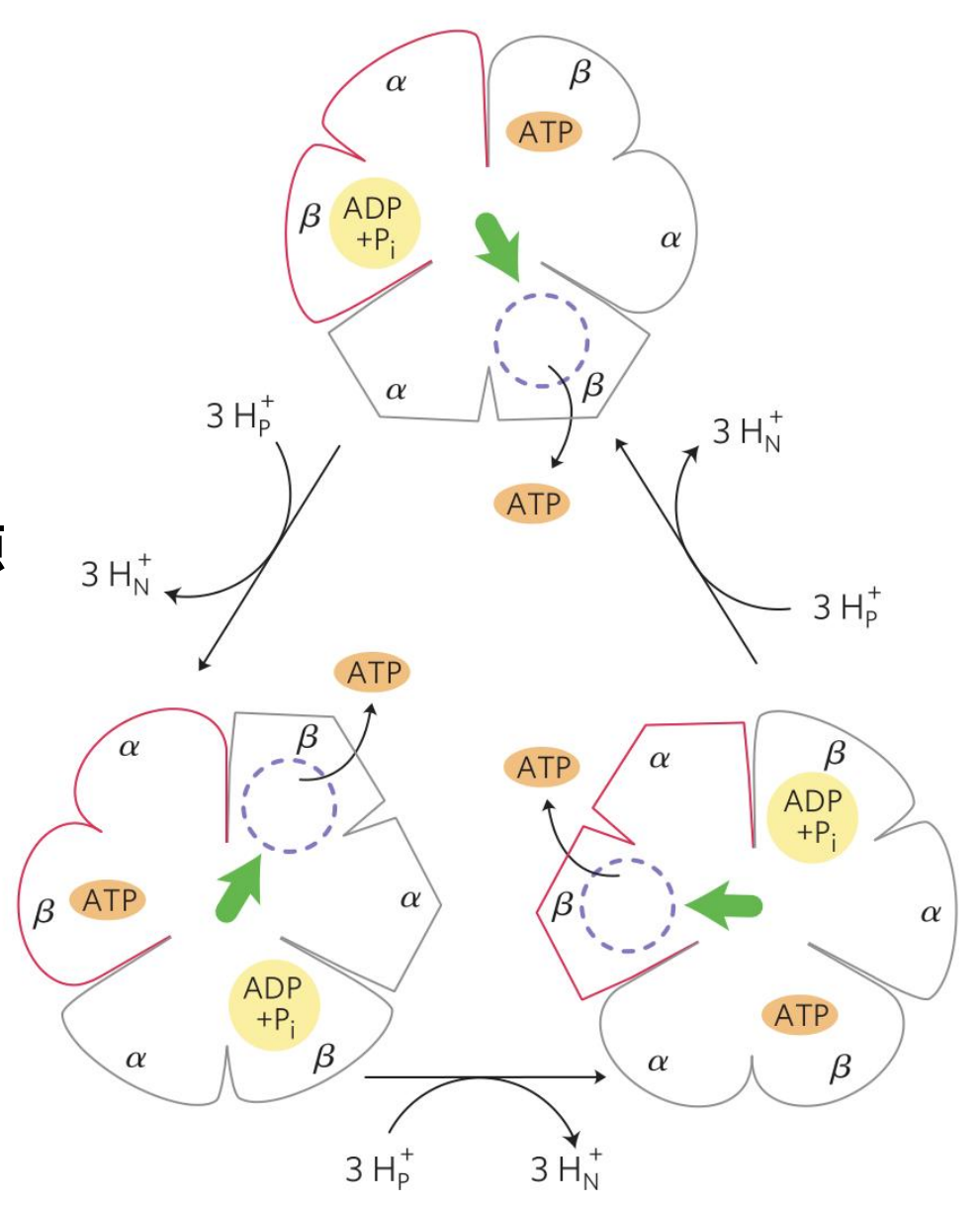
ATP合成酶的中杆是不对成的,其作为“传动杆”驱动三个“ATP合成车间”——\(\alpha\beta\)异源二聚体按顺序合成ATP。每转动一周,它就会按顺序“挤压”三个异源二聚体,使得其采取三种不同的开放构象:
- 最紧密的是\(\alpha/\beta_{\rm{DP}}\)
- 比较紧密的是\(\alpha/\beta_{\rm{TP}}\)
- 最开放的是\(\alpha/\beta_{\rm{E(empty)}}\)