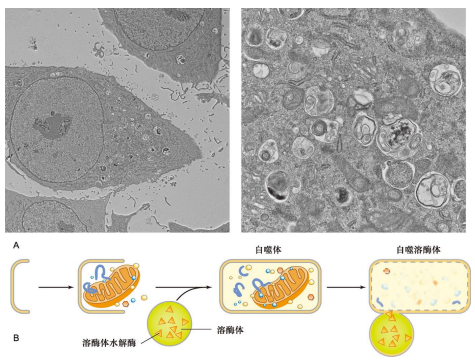
细胞衰老与死亡
细胞的增殖、衰老与死亡也是保持生物稳态 (Homeostasis) 的一环。
细胞衰老 Cellular Senescence
细胞衰老是指随着时间的推移,细胞增殖能力和生理功能逐渐下降的变化过程。
细胞衰老具有如下生物学表现:
- 细胞周期抑制 (Cell cycle arrest)
- 细胞的形态学改变 (Morphological transformation)
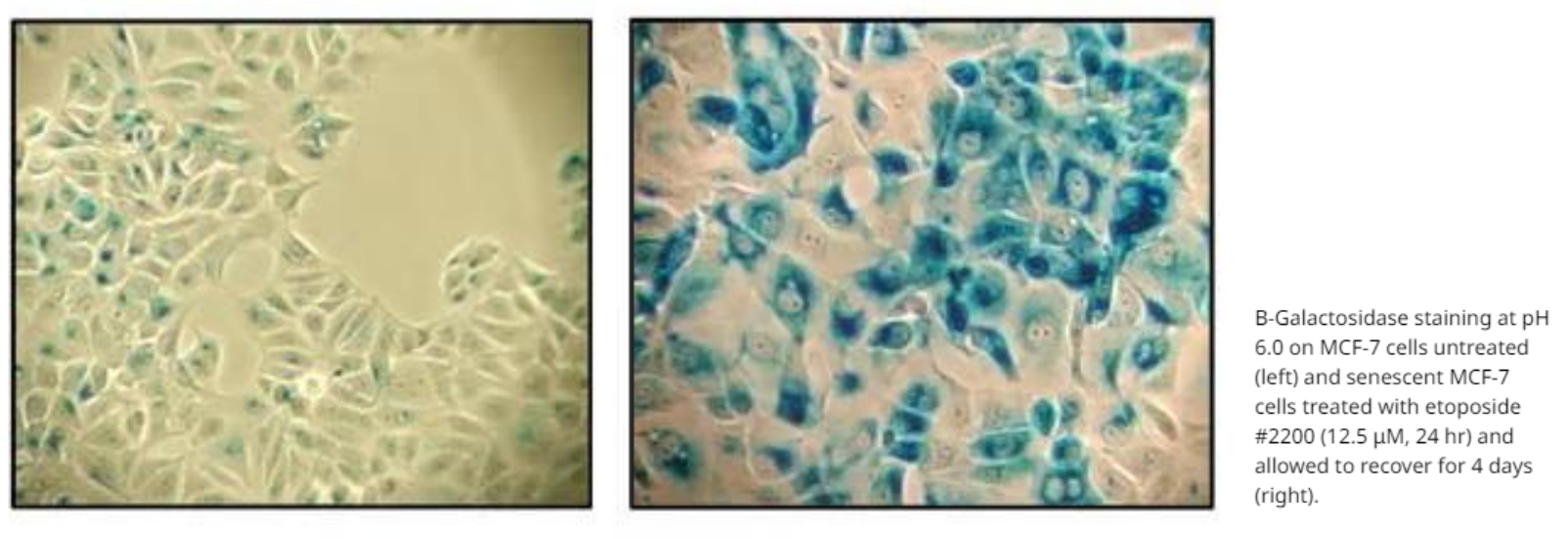
- β-半乳糖苷酶 (SA-β-GAL) 的活化
- 衰老相关异染色质聚集 (Senescence-associated heterochromatic foci, SAHF)
- 衰老相关分泌 senescence-associated secretory phenotype, SASP)
衰老细胞发生一系列形态学上的改变,细胞内β−半乳糖苷酶发生活化。
细胞衰老类型
细胞有如下两种衰老类型:
- 细胞的复制衰老 (Replicative senescence) —— 寿终正寝
- 细胞的胁迫诱导衰老 (Stress-induced premature senescence) —— 天灾人祸
复制衰老
细胞的复制衰老主要指体外培养的正常细胞经过有限次数的分裂后,停止生长,细胞形态和生理代谢活动发生显著改变的现象。
Note
1958年,Hayflick等人证实人成纤维细胞的复制能力是有限的,并首次提出了细胞水平上的衰老现象,称为“Hayflick limit”。
他从老年男性与青年女性身上取得人成纤维细胞,实验组为青年女性细胞+老年男性细胞,对照组为老年男性细胞,最终仅有实验组剩下部分细胞,通过观察巴氏小体确认为青年女性细胞,证明了细胞复制衰老的存在,且和环境因素无关。
Hayfilck limit 有两个结论:
- 体外传代的细胞寿命是有限的
- 细胞衰老的过程是由细胞自身决定的,而非环境因素
但是第二点中的后半句话有失偏颇。
细胞寿命
复制衰老的存在告诉我们细胞的寿命是有限的,但是这个限制来自于哪里呢?
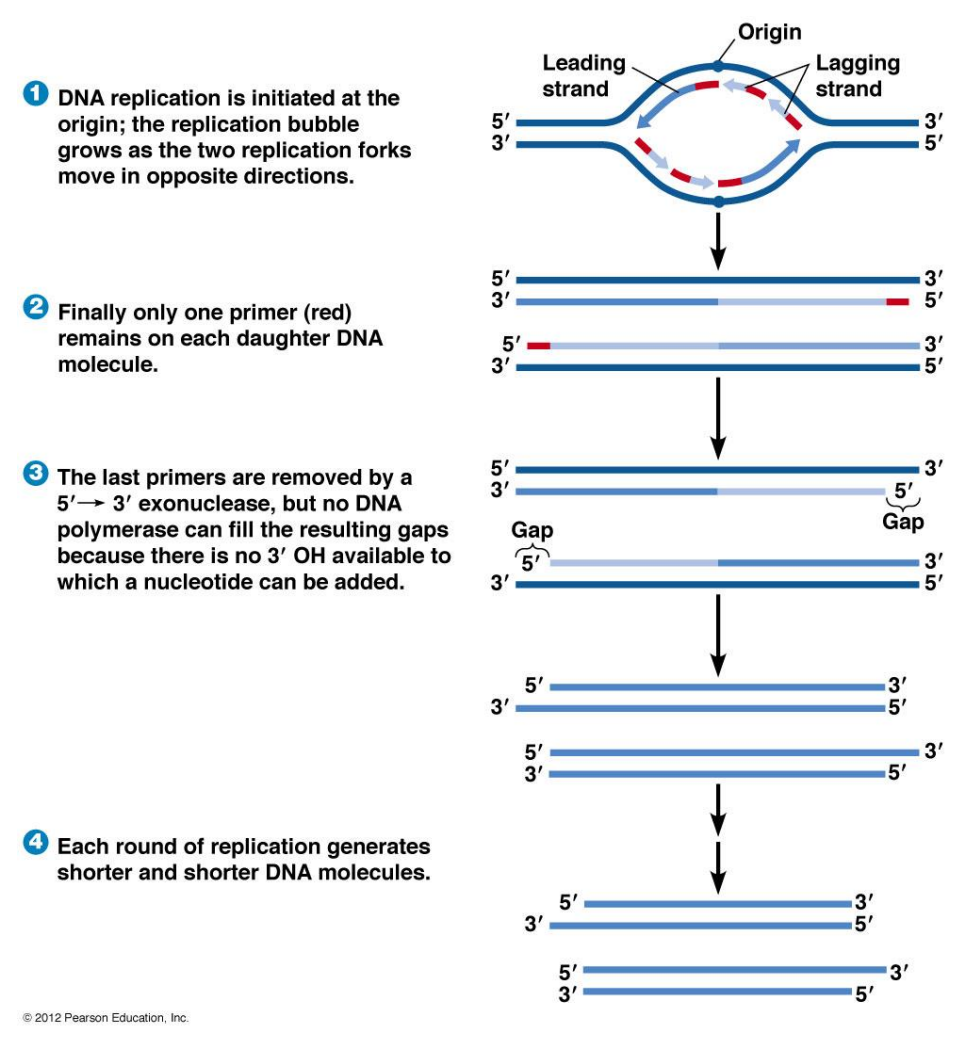
答案是端粒,在细胞核相关章节我们已经了解到了在DNA复制过程中必然出现的末端隐缩问题,端粒就是为了修复这个bug而产生的结构。
端粒
端粒是染色体末端的一种DNA-蛋白质结构,其DNA由简单的串联重复序列组成。人类的染色体端粒DNA由TTAGGG/CCCTAA重复序列组成。
正常情况下在染色体3'末端有大量TTAGGG重复,其会在末端与蛋白形成一个T-loop结构,来保护DNA双链的末端:
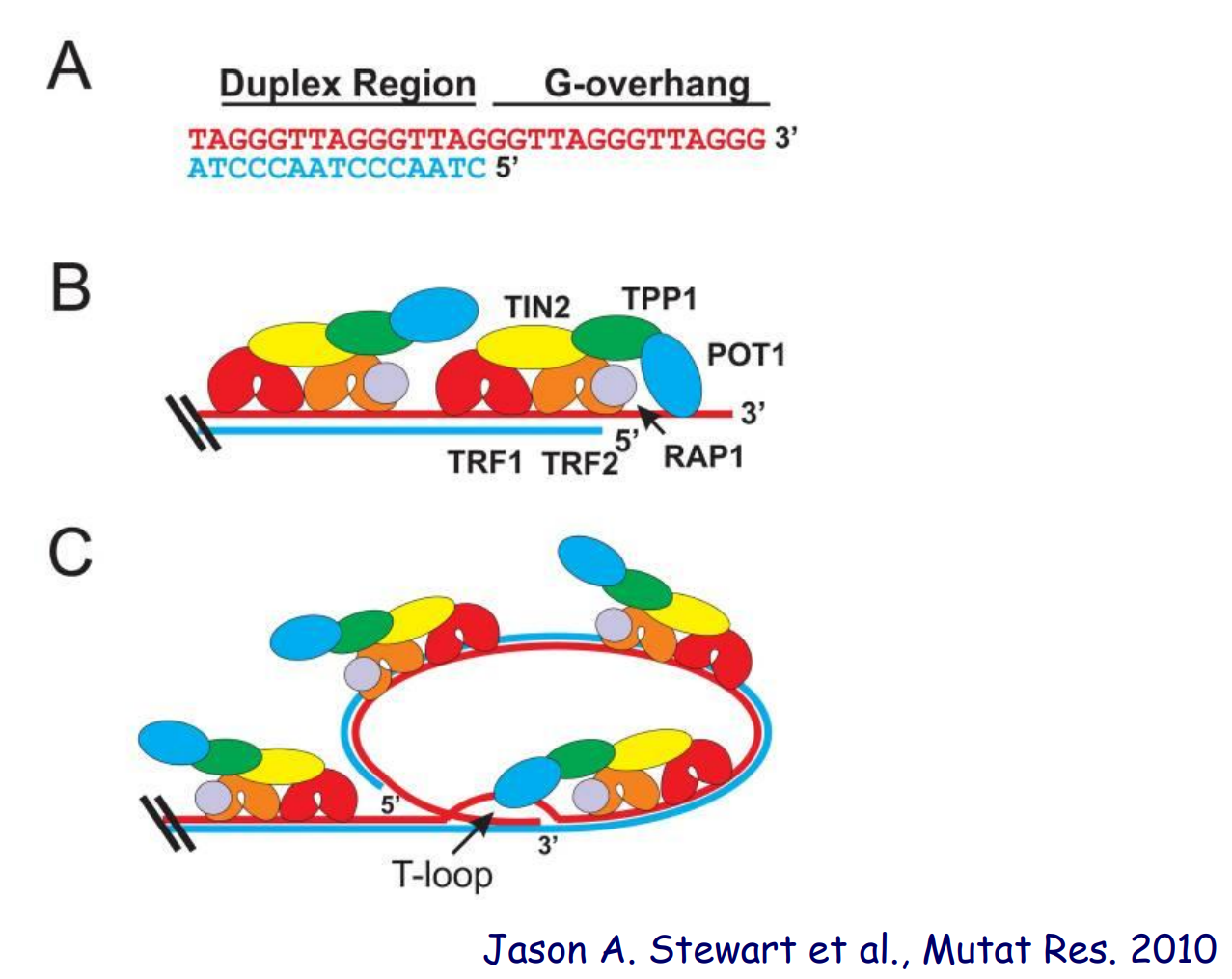
同时端粒还具有另外两个功能:
- 保护染色体不被核酸酶降解
- 防止染色体末端融合
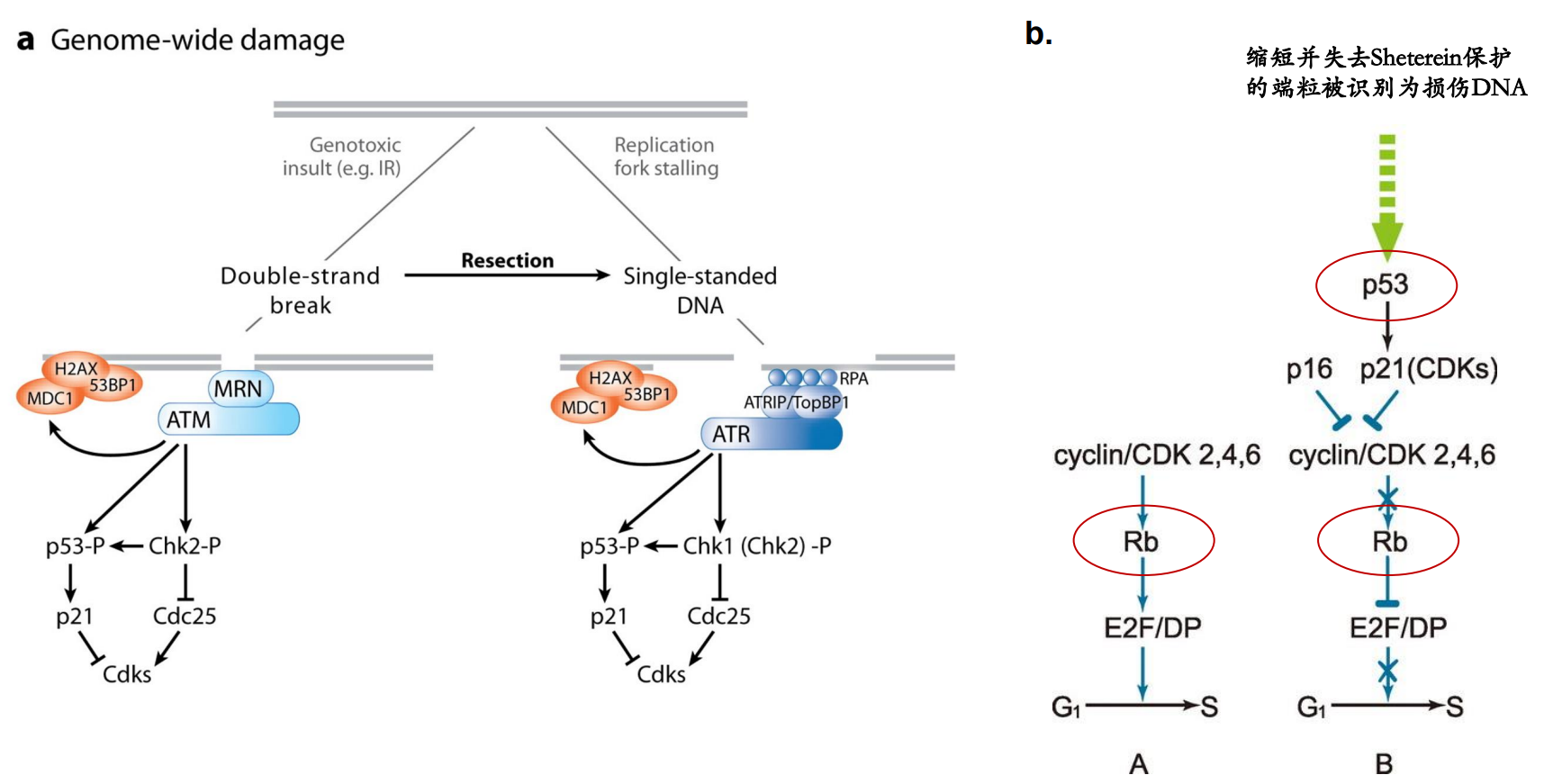
而随着细胞不断的增殖,DNA不断的复制,端粒被不断消耗缩短。当端粒缩短到一定程度时,激活细胞的DNA损伤应答反应 (DNA damage response)。其原理是端粒缩短后 T-loop 结构无法维持,失去了 Shelterin 蛋白的保护的端粒暴露出双端DNA序列,被细胞识别为DNA双链断裂,启动DNA损伤应答。
端粒酶 Telomerase
但是如果不解决端粒问题,所有细胞都有分裂的终点。但是显然对于我们身体里的某些细胞,比如干细胞、癌细胞,它们似乎有无限的分裂潜力。那端粒的限制去哪里了?
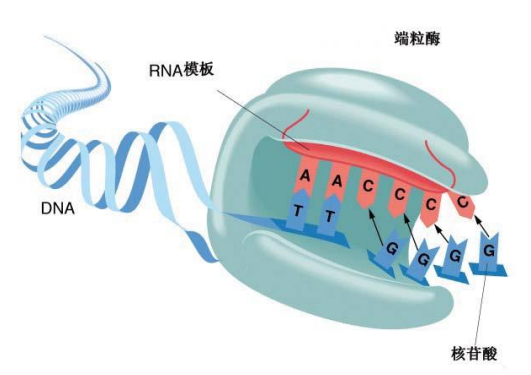
答案是端粒酶,细胞内最专业的端粒修复专家。
端粒酶的生物学实质是由逆转录酶 ( Reverse transcriptase )与RNA组成的核糖核酸-蛋白复合物。它利用自身携带的RNA为模板,催化端粒DNA的合成。
细胞胁迫诱导衰老
细胞的SIPS是细胞在多种亚致死的外因影响下产生的细胞衰老状态。这些因素包括氧化损伤、UV照射等等,但与引起复制衰老的端粒自发性缩短无关。
特别注意的是,SIPS与复制衰老具有相同的表型(形状,β-半乳糖苷酶活化),但是进程更加迅速。
细胞的复制衰老与胁迫诱导衰老都可以视作是有机体在长期演化过程中形成的防止细胞过度生长即癌化的一种机制。但至今没有研究能够关联体外细胞培养的衰老与个体衰老的关系。
细胞死亡 Cellular Death
细胞死亡的类型分类
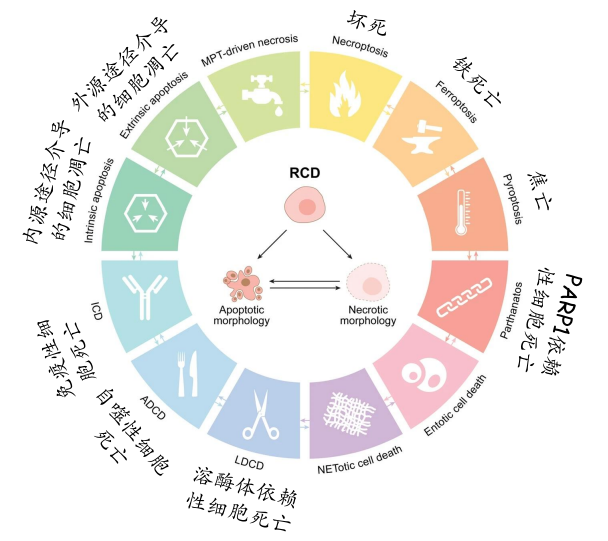
而我们常用的主要是以下三类,也是研究比较多的三类:
- 细胞凋亡 (Apoptosis)
- 细胞坏死 (Necrosis)
- 自噬性细胞死亡 (Autophagic cell death)
细胞凋亡 Apoptosis
细胞凋亡是受基因控制的主动的细胞生理性自杀或谋杀行为,是正常的生理过程。
细胞凋亡特征
细胞凋亡的过程包括:凋亡的起始、凋亡小体的形成与吞噬。
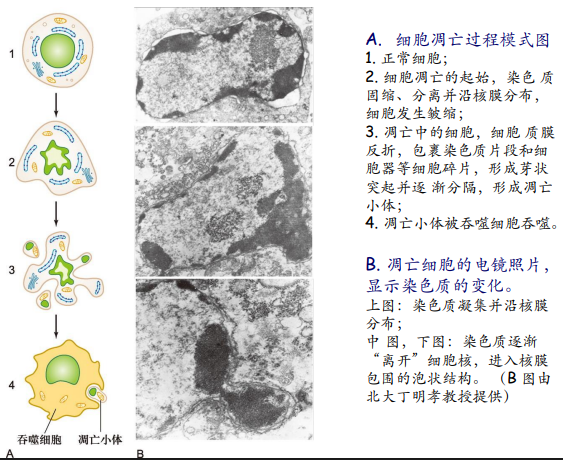
细胞凋亡检测方法
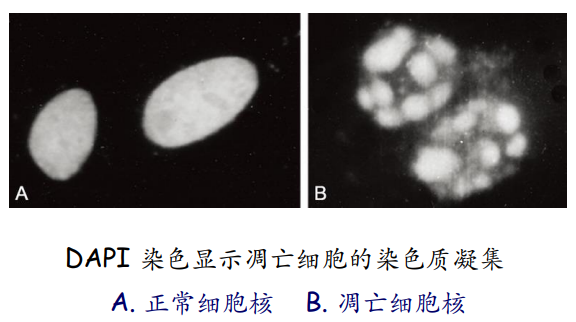
第一种是染色法——台酚蓝、DAPI、吉姆萨染色法 (Giemsa)
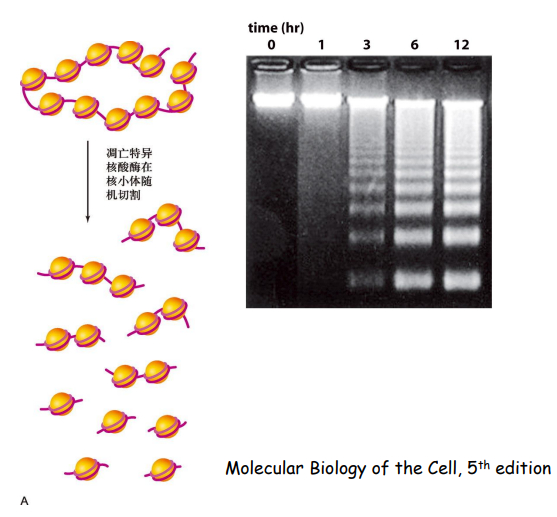
第二种是 DNA 电泳(金标准),凋亡时,细胞内特异性核酸内切酶活化,染色质DNA 在核小体间被特异性切割,DNA 降解成 180~200 bp或其整数倍片段。
其结果是在细胞的DNA电泳结果中形成DNA梯状条带 (DNA ladders)。
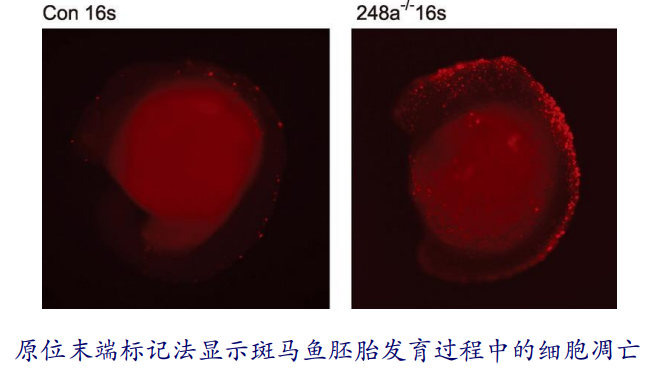
第三种是 DNA 断裂的原位末端标记法。凋亡细胞的核DNA 断裂后产生3′-OH 末端,因此可借助一种可观测的标记物如荧光素对单个凋亡细胞核或凋亡小体进行原位染色。
比如暴露的3-OH在末端脱氧核苷酸转移酶(Terminal Deoxynucleotidyl Transferase, TdT)的催化下加上荧光素标记的dUTP(fluorescein-dUTP),完成原位标记。
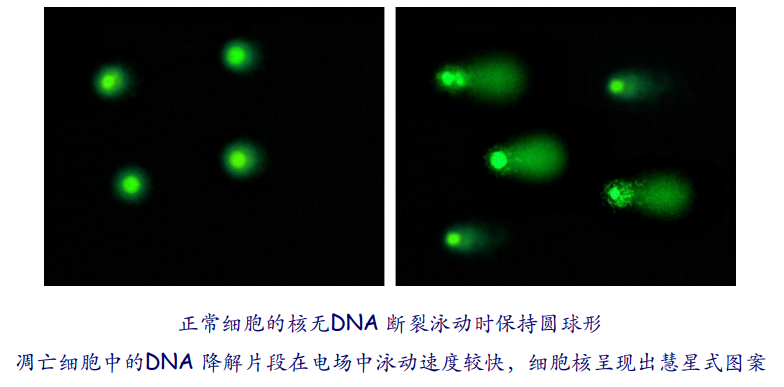
第四种是彗星电泳法,利用染料观察电泳中细胞的DNA总体形态。
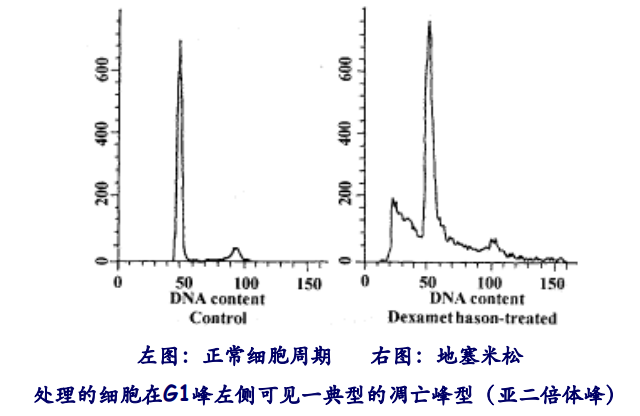
第五种是流式细胞术,将细胞用DNA染料染色后用流式细胞仪来测定DNA含量。(形成高峰左侧的sub G1期,因为DNA随着凋亡小体被丢出了细胞)
第六种检测细胞的细胞膜成分,最经典的是检测磷脂酰丝氨酸的外翻 (eat-me signal),此外还有caspases激活、Cyt C释放、线粒体膜电位变化等等。
细胞凋亡早期,位于细胞膜内侧的磷脂酰丝氨酸(PS)迅速翻转到细胞膜外侧,使得PS 暴露在细胞膜表面,可以用PS 与抗凝血剂Annexin V 的相互作用检测外翻的PS。
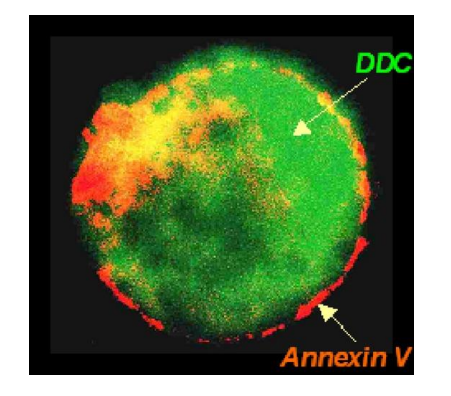
下图展示了细胞如何利用 PS 外翻来传递 eat-me 信号。
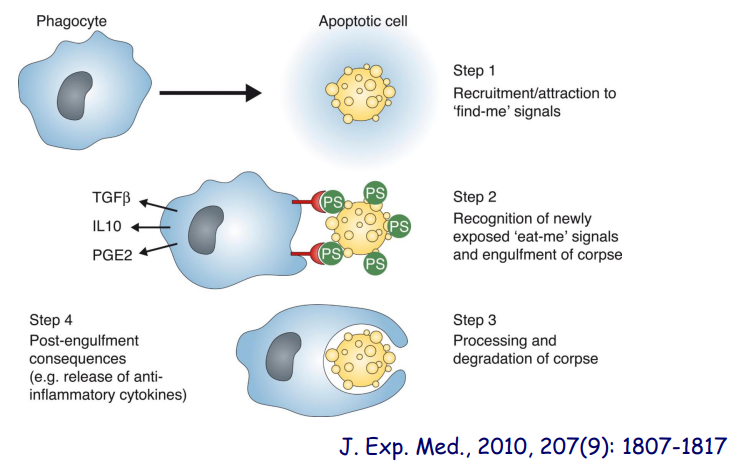
细胞凋亡分子机制
动物细胞的凋亡阶段分为:
- 接受凋亡信号
- 凋亡相关分子活化
- 凋亡执行
- 凋亡细胞清除
其中动物细胞推动以上凋亡过程主要有两条途径:Caspase依赖性和不依赖于Caspase的细胞凋亡。
caspases
caspases 实际上是 Cysteine aspartic acid specific protease 的缩写,中文名字是天冬氨酸特异性的半胱氨酸蛋白水解酶。 是一组存在于细胞质中具有类似结构的蛋白酶,活性位点均包含半胱氨酸残基,能够特异地切割靶蛋白天冬氨酸残基后的肽键。
caspases 的职责是选择性切割某些蛋白质,使其失活或者活化,但是并不是降解。
一般情况下,caspases 会以无活性酶原形式存在于细胞质中。
我们主要介绍Caspases依赖性的细胞凋亡。
在途径中,一开始是Caspases的激活,通过同性活化与异性活化两种方式,依靠级联效应启动凋亡程序。
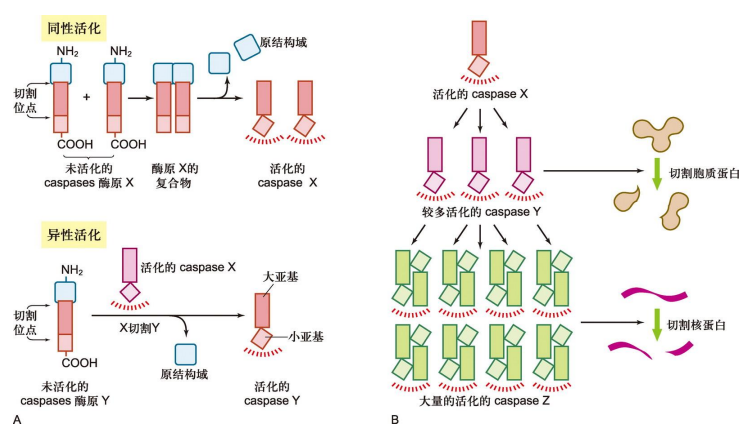
之后 Caspases 需要切割一系列下游靶蛋白,使得细胞呈现出凋亡的一系列形态学和分子生物学特征,它们包括:
- 切割后被活化的蛋白,其代表就是核酸酶 CAD
- 切割后失活的蛋白,这些蛋白一般在维护细胞正常状态中发挥重要作用。
- 细胞骨架蛋白被切割后使得细胞形态发生改变,有利于凋亡小体的形成。
Caspases 依赖性细胞凋亡途径的上游起始有两个:
- 死亡受体起始的外源途径
- 起始于死亡配体与受体的结合
- 死亡配体主要是肿瘤坏死因子(TNF)家族成员;胞质部分均含有死亡结构 域,负责招募凋亡信号通路中的信号分子
- 线粒体起始的内源途径
- 内源途径中,线粒体处于中心地位
- 当细胞受内外部凋亡信号刺激时,胞内线粒体的外膜通透性会发生改变,向细胞质中释放凋亡相关因子,引发细胞凋亡
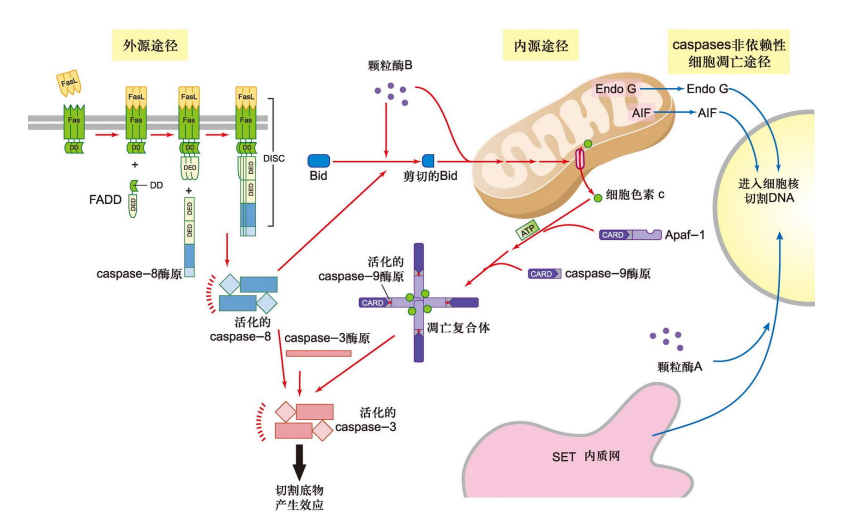
Caspases 非依赖性细胞凋亡途径我们简略通过一张图片了解一下,其通过凋亡诱导因子和限制性内切核酸酶 G诱导。
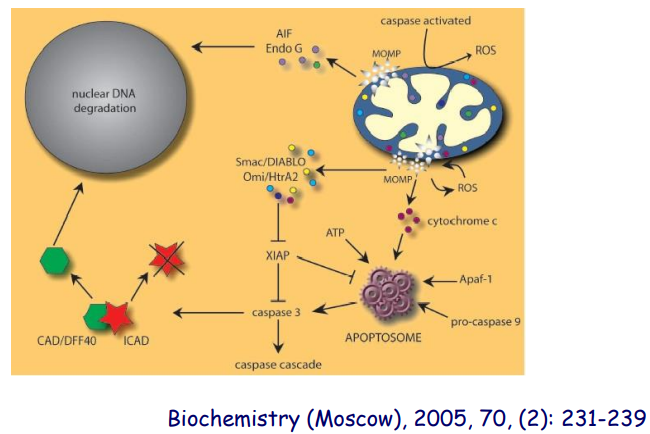
细胞凋亡的调控
仅作了解:生存因子通过调节Bcl-2家族成员的活性及表达抑制细胞凋亡的发生
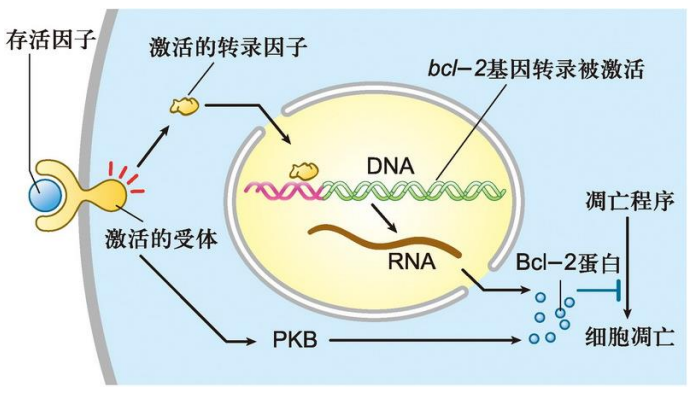
Bcl-2 家族

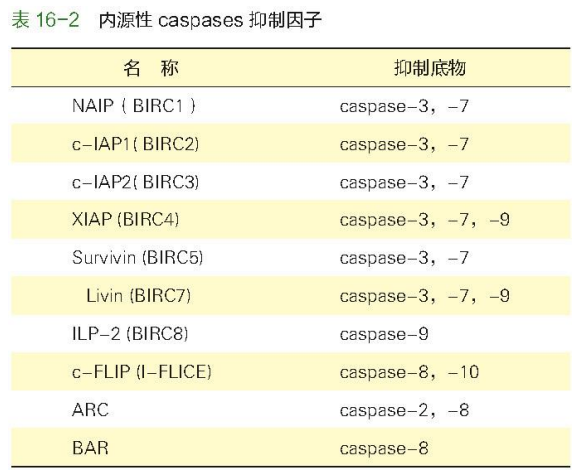
下图展示了 Caspases 抑制因子:
细胞凋亡的生理学与医学意义
- 细胞凋亡在正常发育、自稳态的维持、免疫耐受的形成、肿瘤监控等过程中均发挥重要作用:
- 确保正常发育、生长
- 维持内环境稳定
- 发挥积极的防御功能
- 凋亡失调:不恰当激活或抑制;过度凋亡导致疾病
- AIDS:HIV诱导淋巴细胞凋亡
- 自身免疫疾病:激活的淋巴细胞凋亡缺陷
- 癌症:凋亡缺陷
- 老年痴呆/中风:过多神经细胞凋亡
一个典型的例子是神经系统与靶细胞的一一配对:
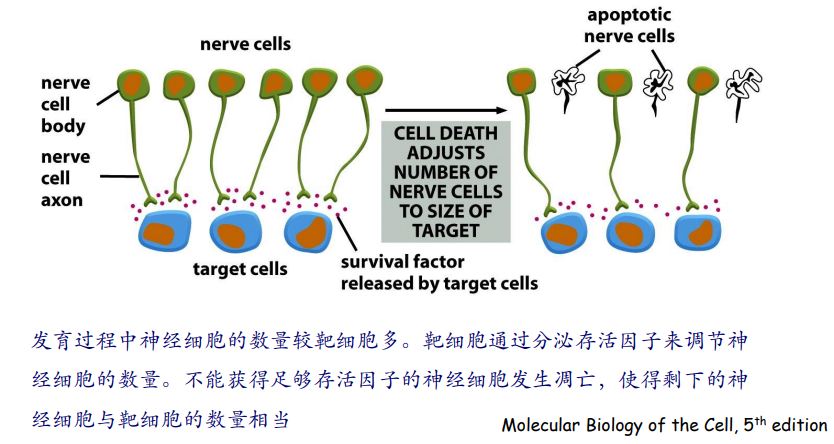
在动物发育过程中,也会发生细胞凋亡的过程:
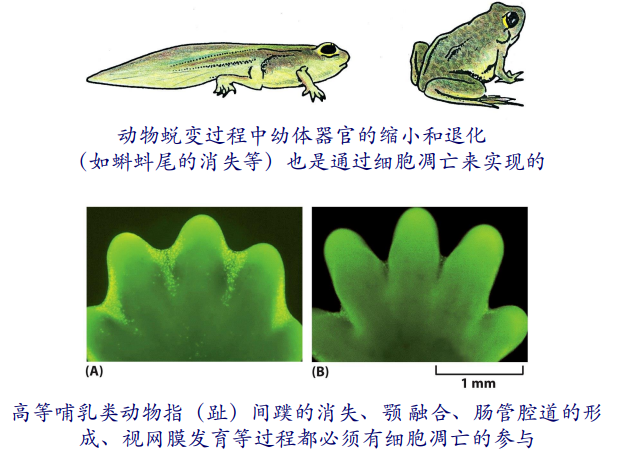
细胞凋亡还在免疫耐受的形成中发挥重要作用:
Note
2025 的诺贝尔生理学与医学奖!
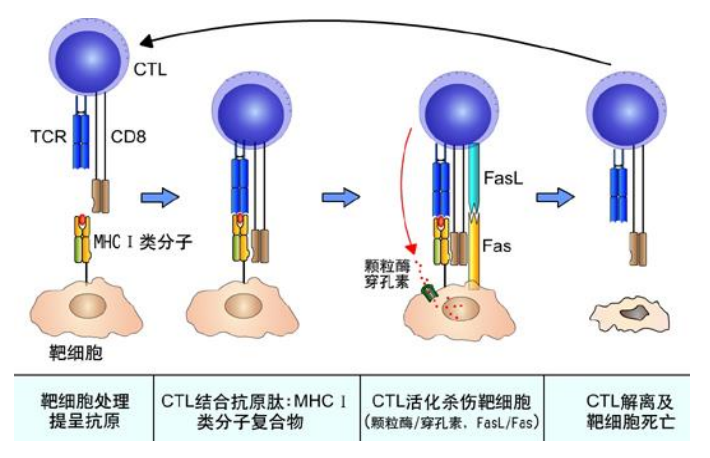
细胞凋亡让免疫系统可以剔除被病原体感染的细胞:
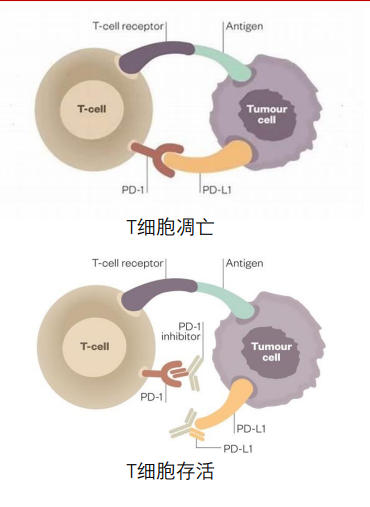
针对PD1与PD-L1的抗体的肿瘤治疗策略也与细胞凋亡有关。
Programmed Death 1,简称PD1,是表达于T细胞表面的受体。某些肿瘤细胞通过表达PD-L1(PD1配体)与PD1结合,诱导T细胞发生凋亡,从而完成免疫逃逸。
通过利用 PD1 抗体和 PD-L1 抗体阻断肿瘤细胞对 T 细胞的凋亡诱导,均能够起到激活免疫系统对肿瘤细胞的识别及杀伤作用。
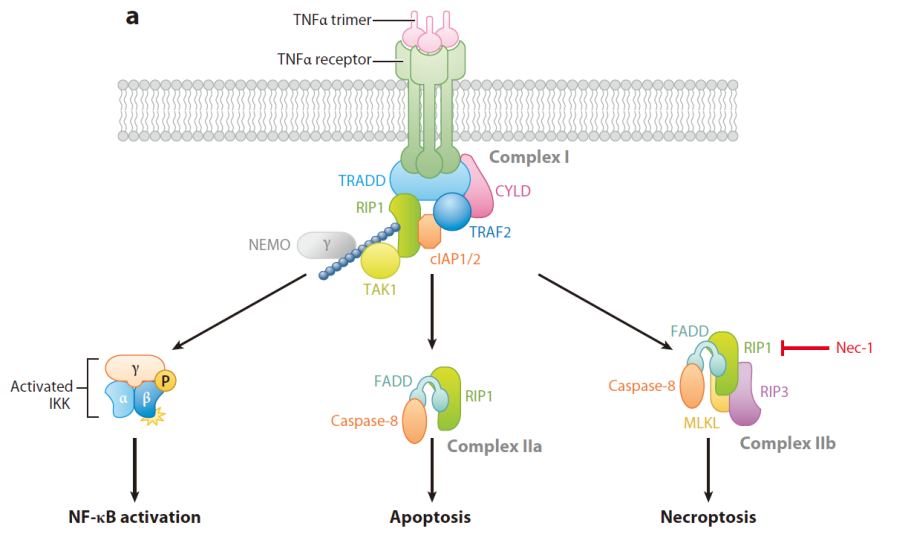
细胞坏死 Necrosis
下图对比了坏死和凋亡的区别:
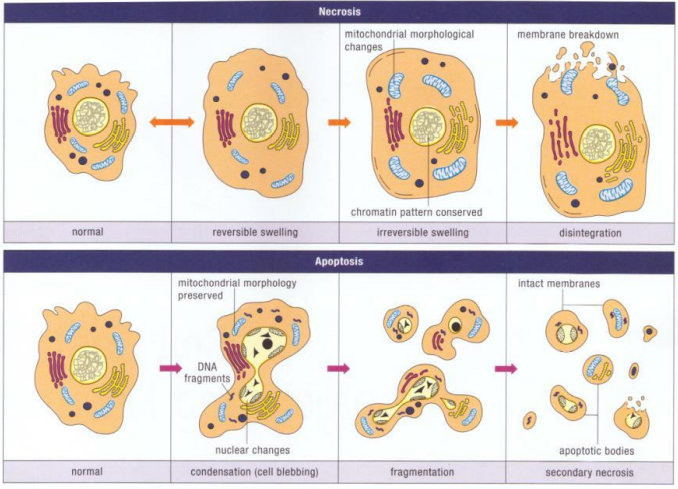
可以看出细胞坏死的过程更加剧烈,不形成凋亡小体,内容会被直接释放。
新研究表明细胞坏死也受到基因的调控!
下图展示了同一个信号上游受体导致的三种不同的细胞命运:
细胞自噬性死亡 Autophagic cell death
细胞自噬普遍存在于真核细胞中,它以溶酶体对细胞自身结构进行吞噬和降解,促进细胞组分再循环。
细胞自噬能清除降解受损伤的细胞结构、衰老的细胞器、以及不再需要的生物大分子等,在抵御病原微生物入侵方面也有重要作用。但是,当细胞自噬现象特别严重时,会引起细胞死亡,这种死亡方式称为细胞自噬性死亡。
细胞自噬性死亡也是程序性死亡的一种。