代谢生化——糖代谢
代谢生化研究的是生物体内具体的生化反应过程,特别是其中的物质变化、能量变化,在正式开始代谢生化内容之前,我们有必要认识生物体内生化反应的几个特点:
- 生物反应遵循热力学定律
- 自发的反应总是向着自由能减小的方向进行(化学原理,即\(\Delta G < 0\)
- 生物体通过ATP实现不同生化反应之间的能量偶联
代谢途径
偶联的系列酶促反应,一个反应的产物是下一个反应的底物,总自由能变化是负数。
新陈代谢对生物的意义有如下几点:
- 从周围环境汲取营养物质
- 将外界营养物质引入为自身结构元件、
- 将结构元件组成自身大分子
- 生命活动所需特殊分子的合成与分解
- 提供生命活动所需的一切能量 (ATP)
代谢生化的研究对象可以被分为两类: 合成代谢与分解代谢。
- 分解代谢途径:以形成ATP的方式从富含能量的分子降解中获得化学能,同时也从中获得NADPH、NADH等还原力,并将食物中的分子转变为合成复杂分子所需的构造单位(结构单元)。
- NADH: 烟酰胺腺嘌呤二核苷酸
- NADPH: 烟酰胺腺嘌呤二核苷酸磷酸
- 合成代谢途径:将小分子合成为复杂分子,其过程需要能量和还原力,分别以ATP和NADPH的形式提供。
代谢生化不仅关注具体的反应过程,还关注反应中的代谢中间产物,仅以糖代谢举例,它就涉及到很多重要的代谢中间产物:6-磷酸葡萄糖、3-磷酸甘油醛、丙酮酸、乙酰辅酶A、草酰乙酸、α-酮戊二酸、谷氨酸与天冬氨酸等等。
糖代谢概况
糖类
糖是富含能量的分子、重要碳源、组织结构的重要成分,也是生物活性物质的构成部分。
糖可以分为:多糖、寡糖、双糖与单糖。
- 多糖:淀粉,纤维素与几丁质等
- 寡糖:3-10个糖分子
- 双糖:蔗糖、麦芽糖、乳糖
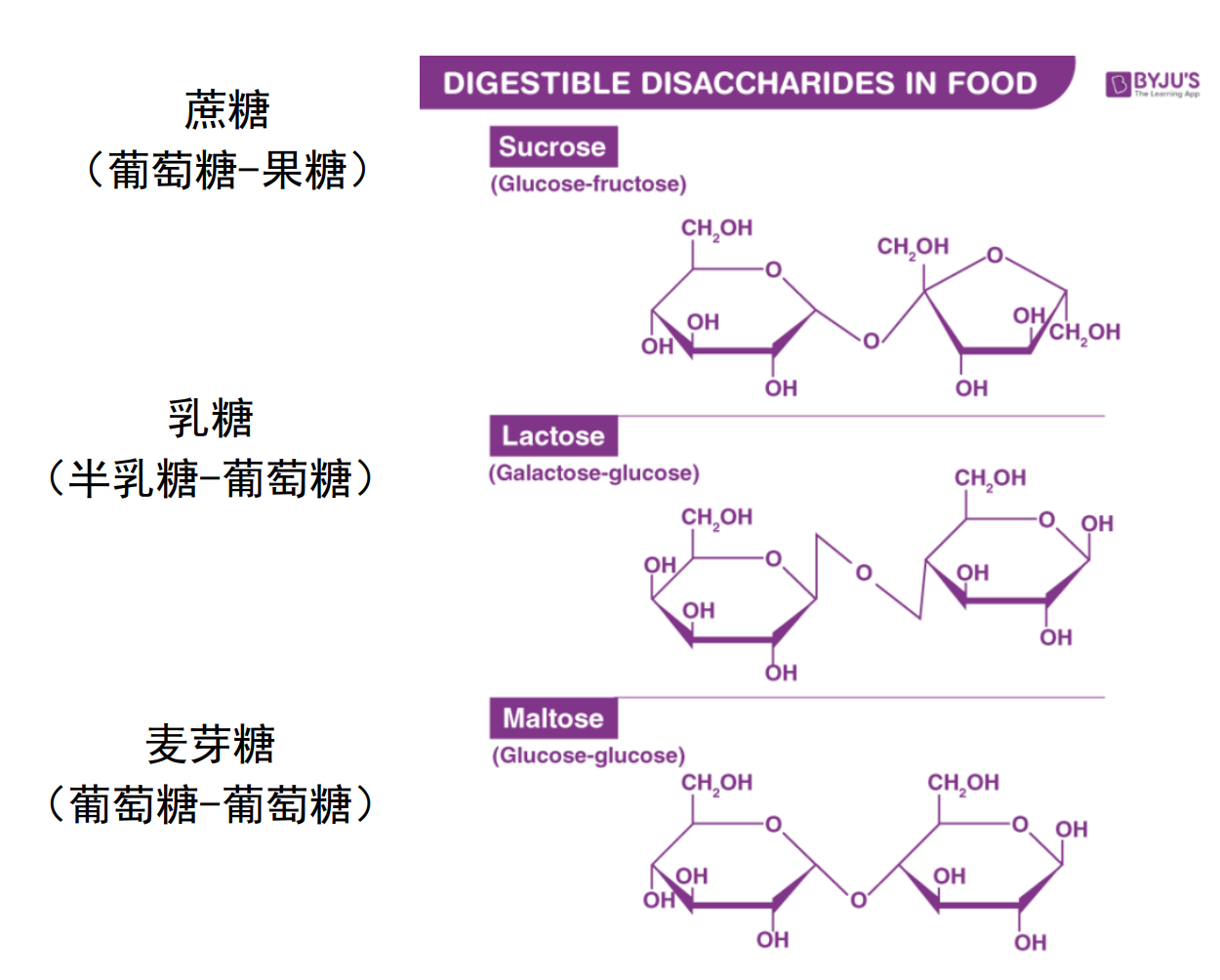
- 单糖:葡萄糖、果糖、半乳糖、甘露糖
生物体内的糖类可以根据其官能团的不同分为酮糖与醛糖:
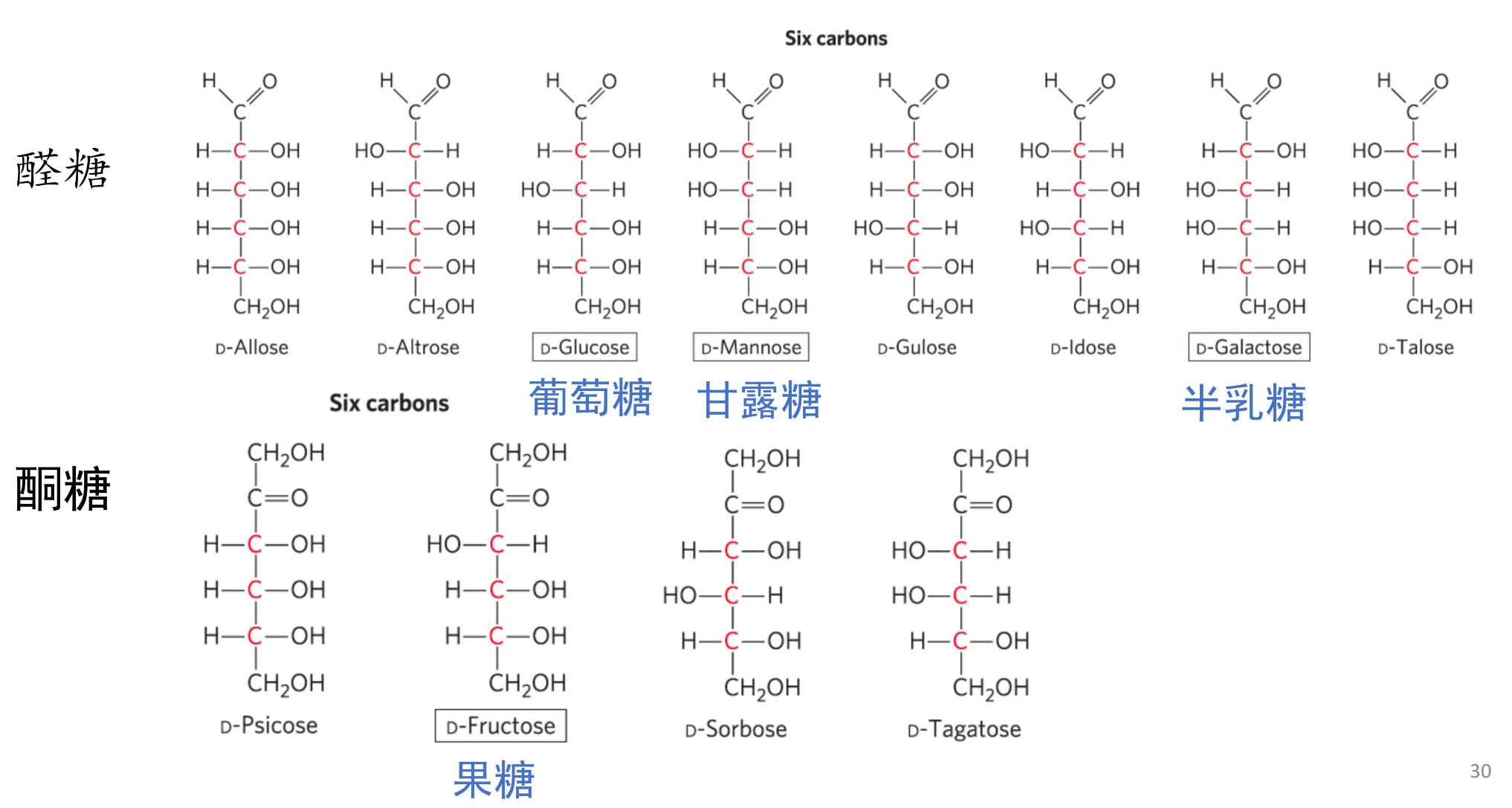
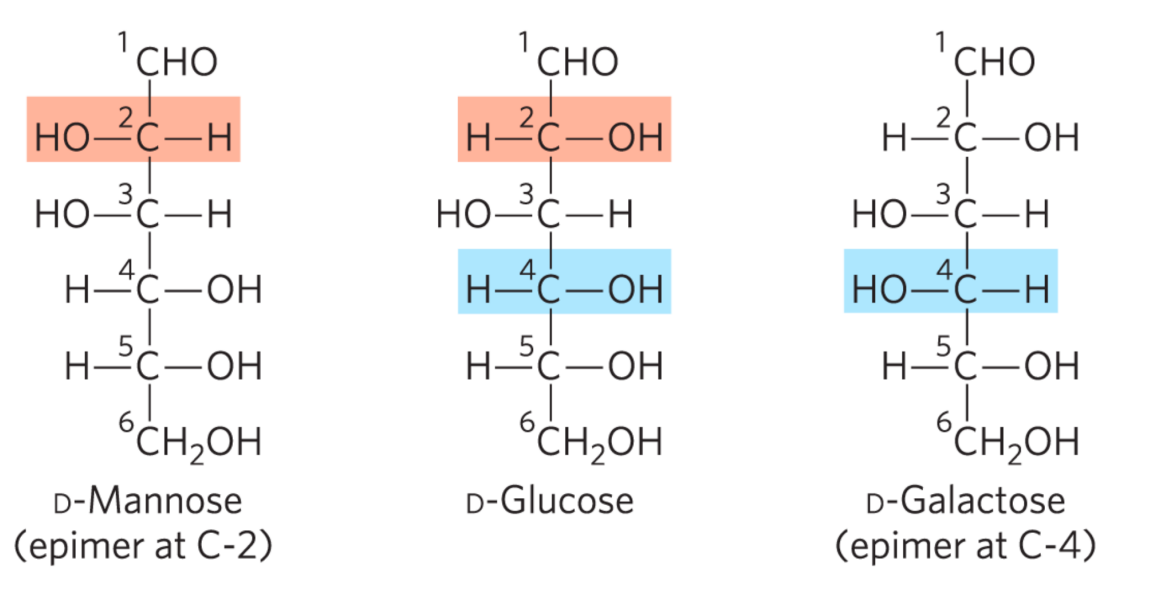
差向异构体
甘露糖和半乳糖都是葡萄糖的差向异构体。
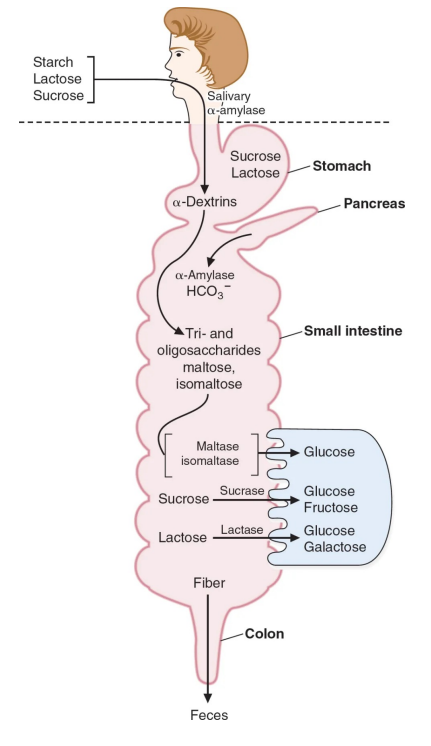
糖的消化与吸收
外界摄入的糖在人体的消化系统内会被逐级消化分解成小分子(单糖),然后被人体吸收利用。
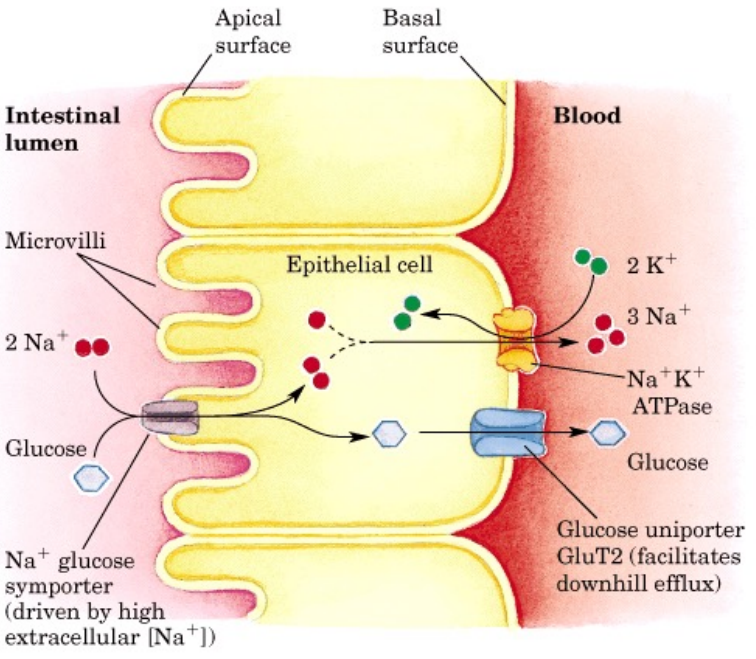
小肠对糖的吸收主要集中在小肠的上半段,小肠上皮细胞通过\(\ce{Na+}\)依赖型葡萄糖转运体吸收葡萄糖,而在肾小管上皮细胞上也有同类的转运体。
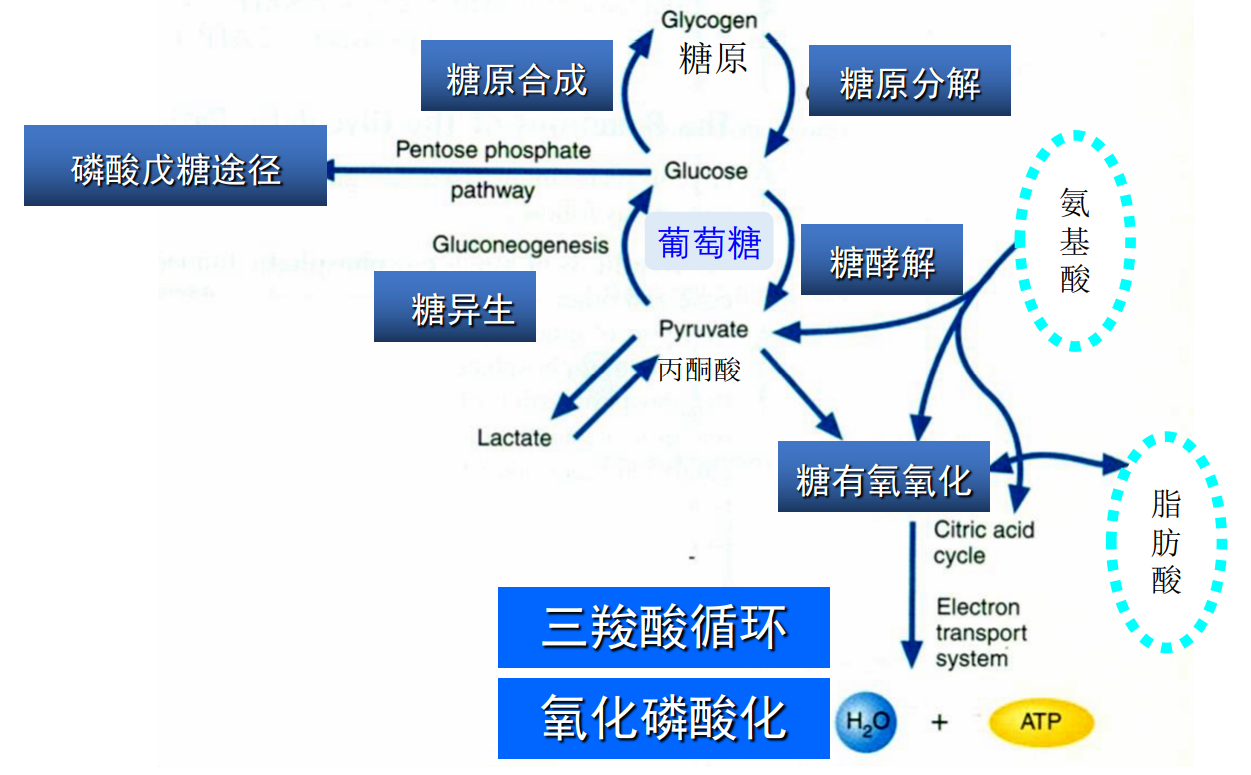
糖的代谢
下图展示了糖代谢的核心内容,葡萄糖的代谢是糖代谢的核心,因为其是哺乳动物大脑等组织非饥饿状态下的唯一能源,是红细胞的唯一能源。血糖稳态对人体的生理稳态至关重要。
糖酵解
糖酵解在不同语境下有不同的含义:
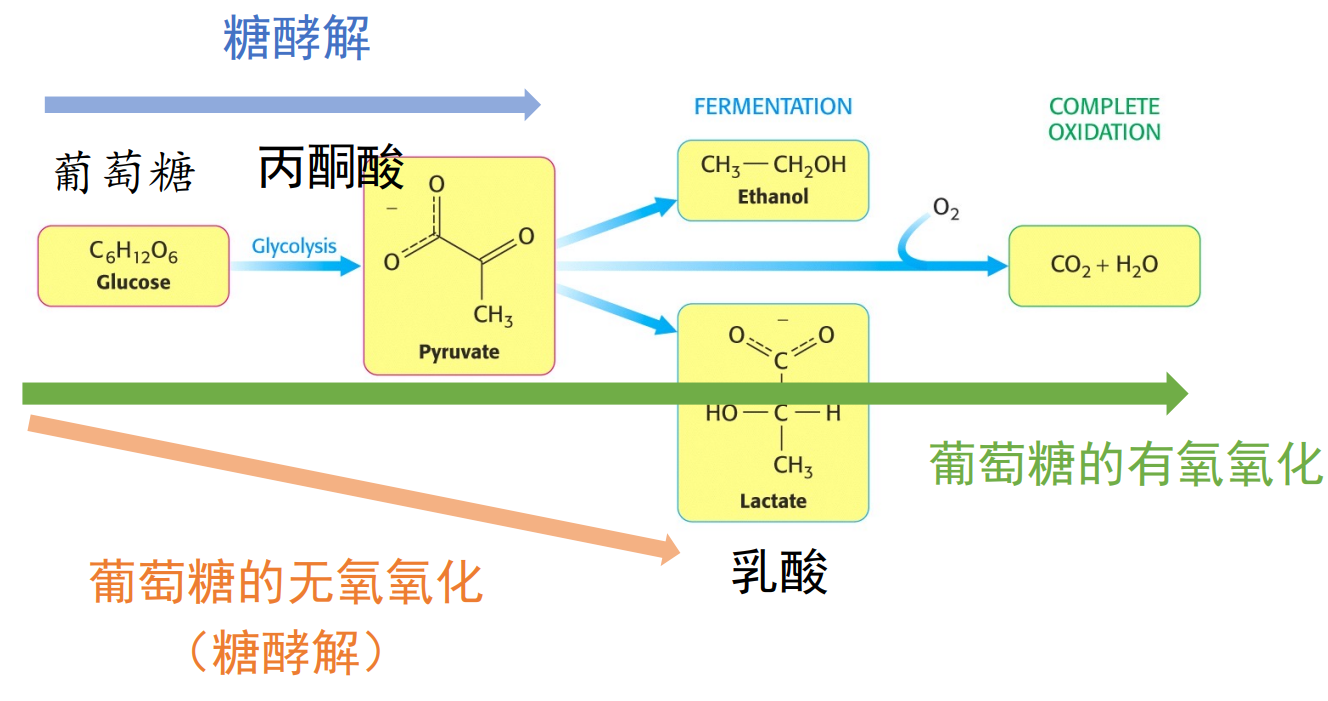
在大多数时候,我们都将糖酵解认为是葡萄糖变为丙酮酸的代谢过程(蓝线),而很多文献也认为葡萄糖的无氧氧化属于糖酵解。
而我们这里讨论的糖酵解遵循前者,是葡萄糖氧化过程中的无氧气参与的阶段。
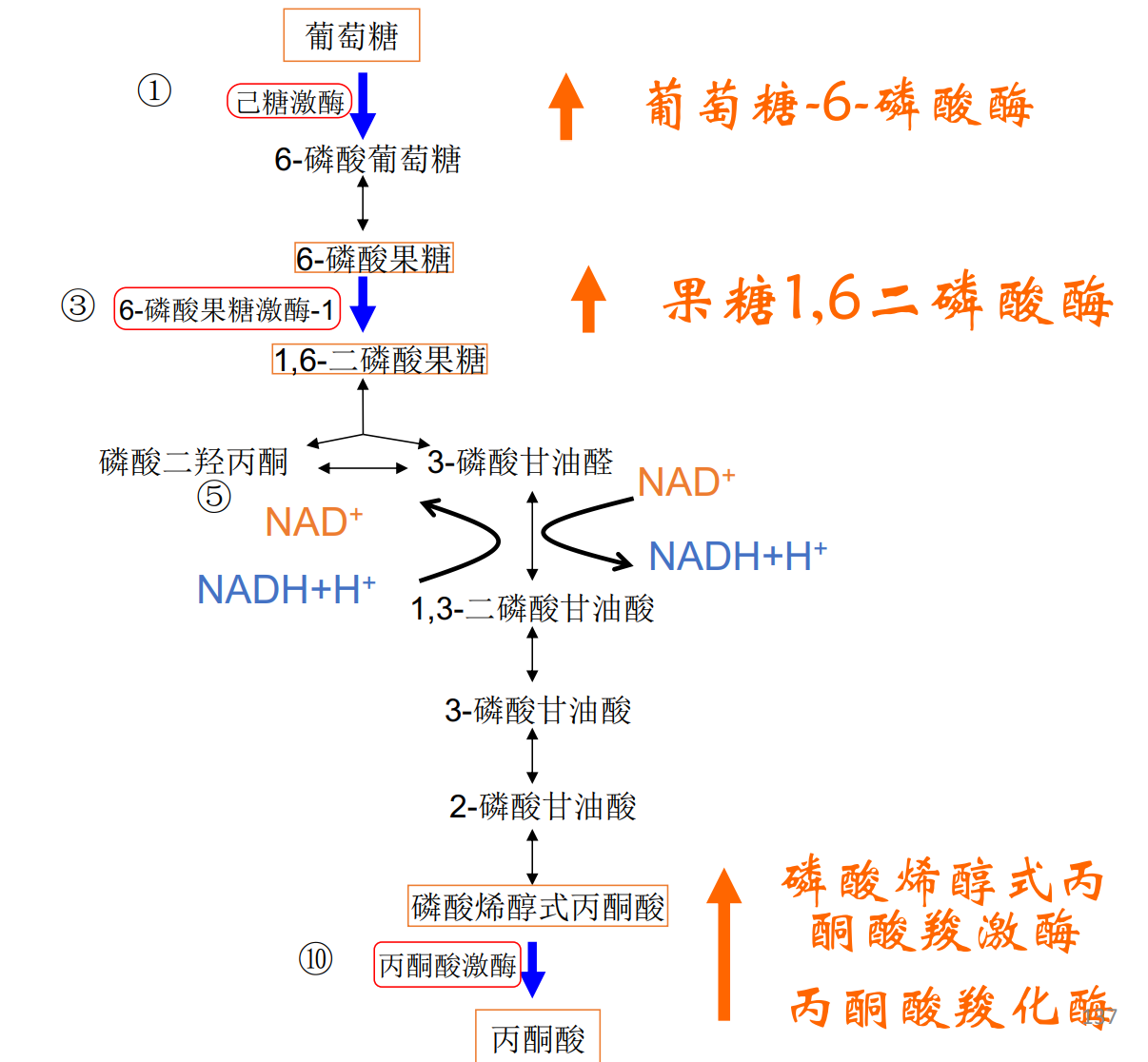
糖酵解是使葡萄糖代谢为两分子丙酮酸并净产生两分子ATP的系列反应,整个过程历经10个步骤,在细胞质基质中进行,整个反应可以分为准备阶段(前五步)与收获阶段(后五步)。
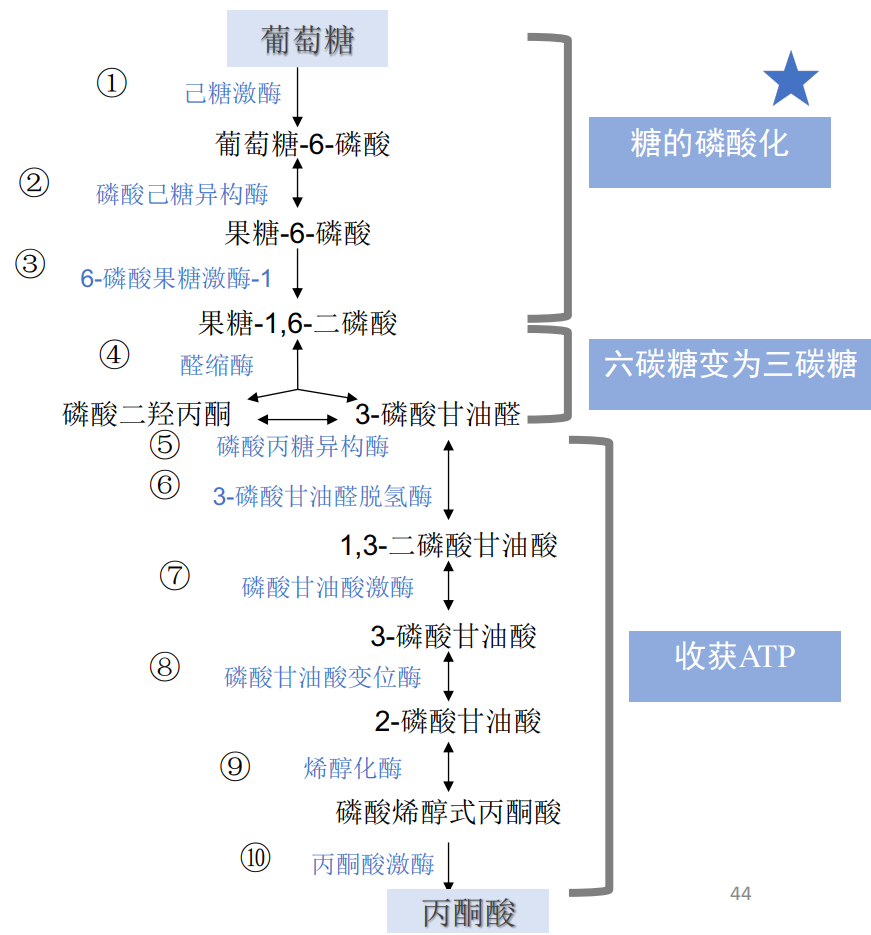
我们接下来按顺序介绍十步反应:
第一步: 己糖激酶磷酸化葡萄糖
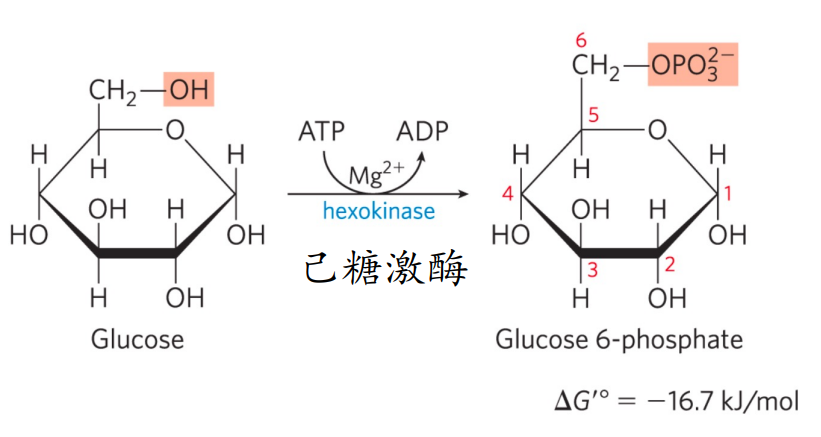
- 磷酸化反应且不可逆。
- 消耗一分子ATP
- 葡萄糖被捕获在细胞内
- 葡萄糖-6-磷酸是重要的结构元件与代谢中间体。
己糖激酶
己糖激酶分为己糖与激酶,前者表示以六碳糖为底物,后者表示从高能分子转移磷酸基团到靶分子的酶。
己糖激酶在非肝脏中还能磷酸化果糖。
与激酶相对的是磷酸酶。
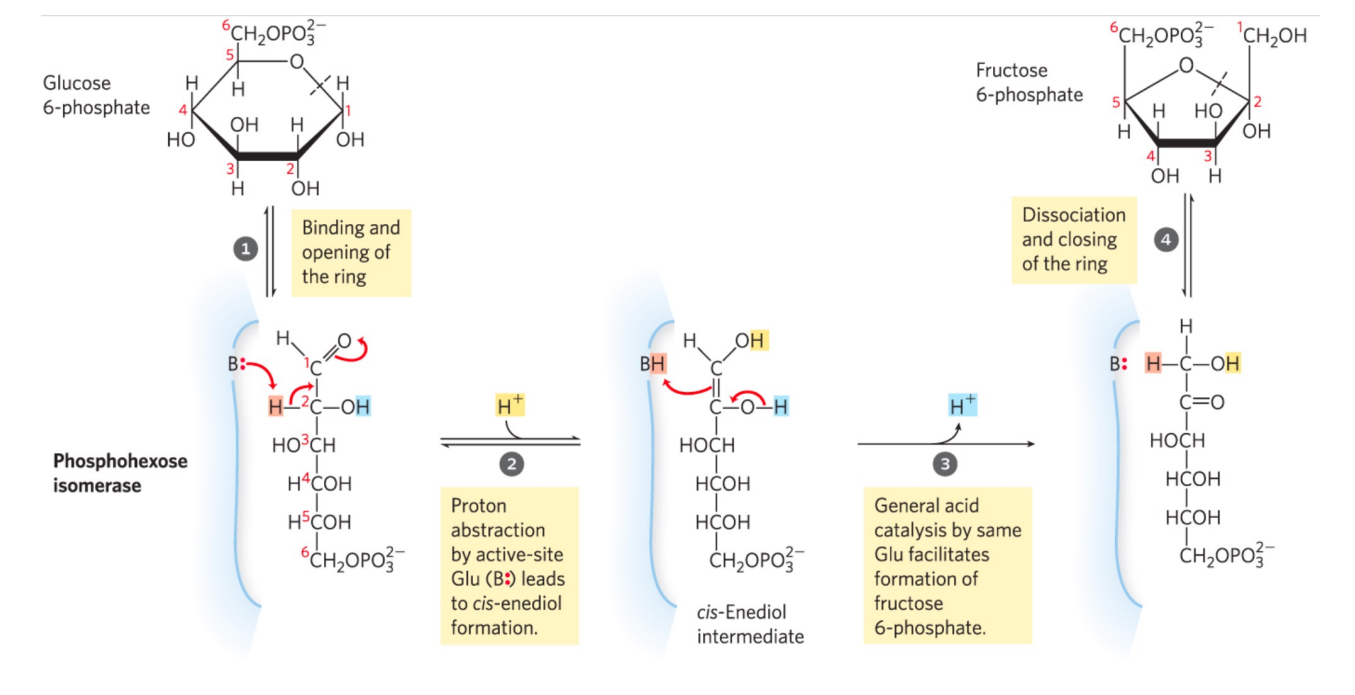
第二步: 磷酸己糖异构酶催化异构
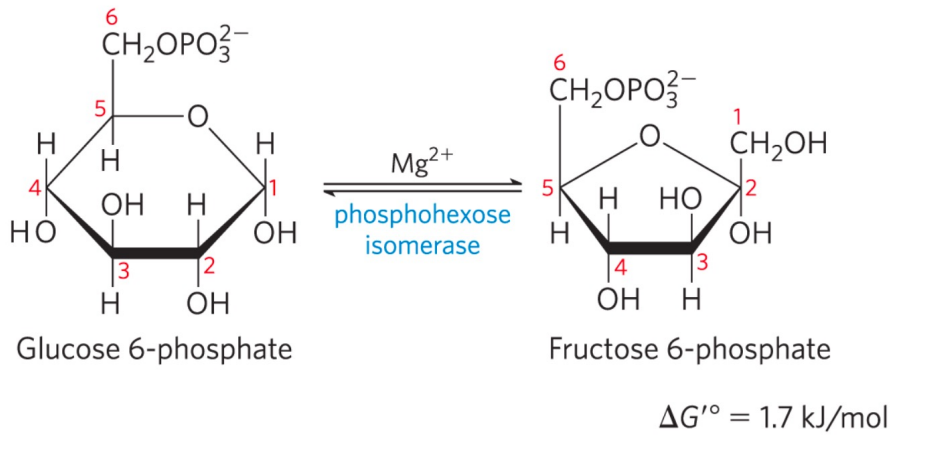
- \(\Delta G \approx 0\),该步可逆,存在两种糖类的转化,但由于下一步不可逆反应的存在,这一步的平衡向右移动。
- 发生分子内重排
磷酸己糖异构酶
磷酸己糖异构酶催化机理如下:
第三步: 6-磷酸果糖激酶-1催化磷酸化
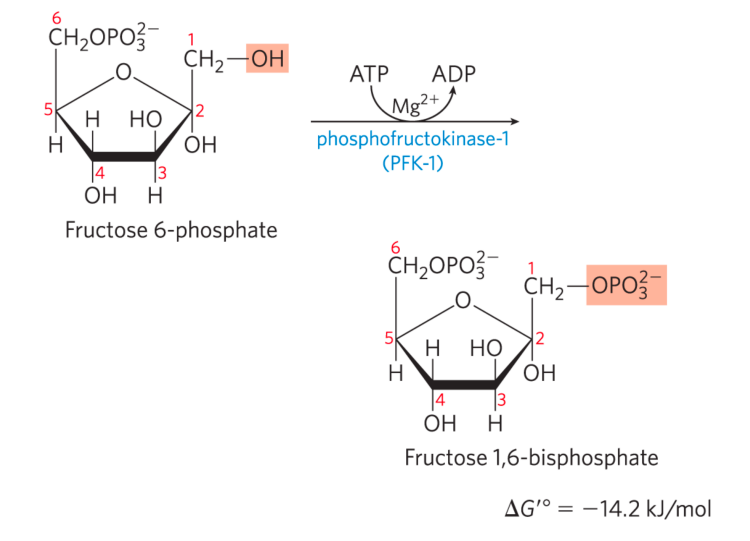
- 该反应不可逆,是确认进入糖酵解的第一步。(相当于守门员)
- 6-磷酸果糖激酶-1,缩写为PFK1,PFK-1与PFK-2不同,后者催化2,6-二磷酸果糖的生成。
- 消耗一分子ATP
第四步: 醛缩酶催化裂解
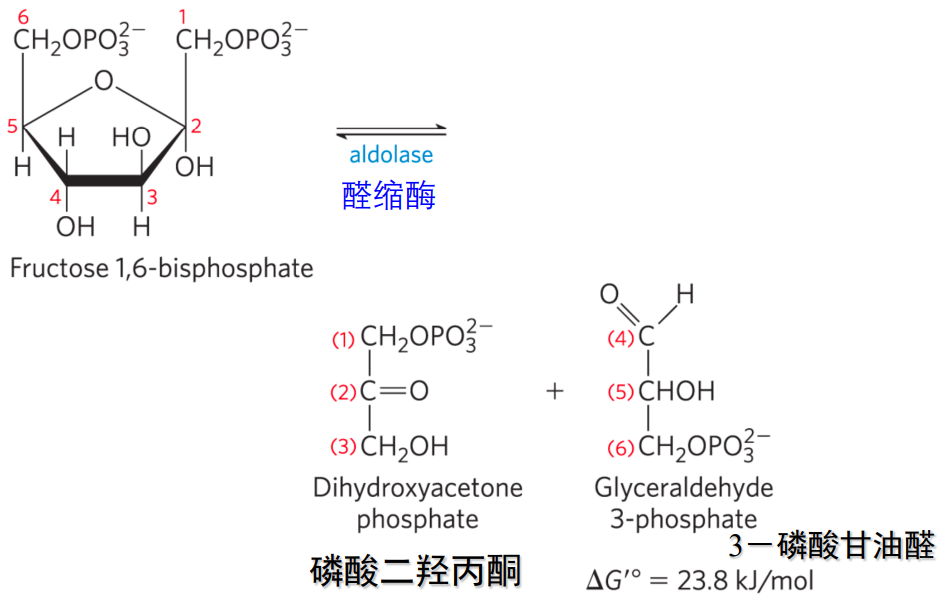
- 本反应是裂解反应,催化此反应的1,6-二磷酸果糖醛缩酶常被简称为醛缩酶。
- 该反应在细胞内基本可逆,\(\Delta G = -1.4kJ/mol\).
该步中碳原子的分配如下:
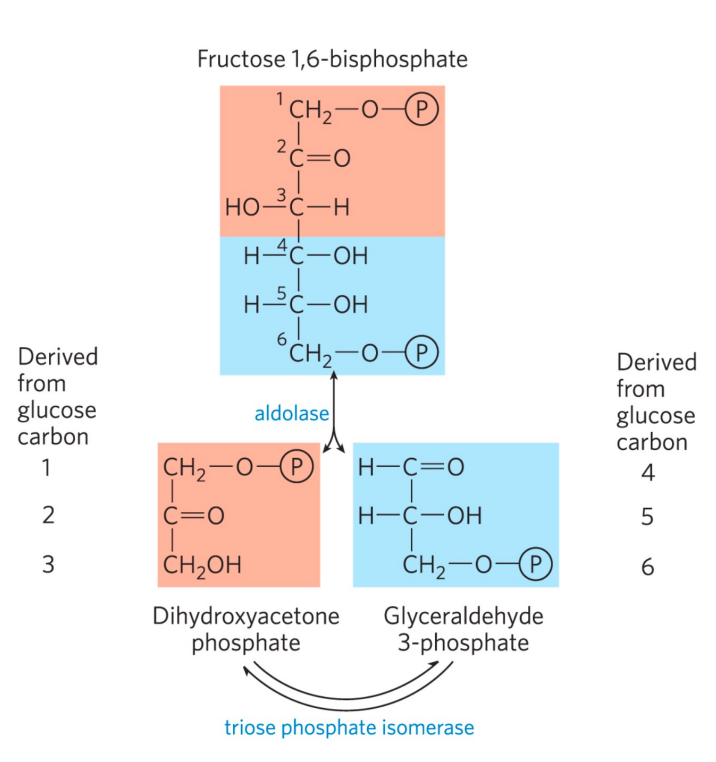
第五步: 磷酸丙糖异构酶催化异构
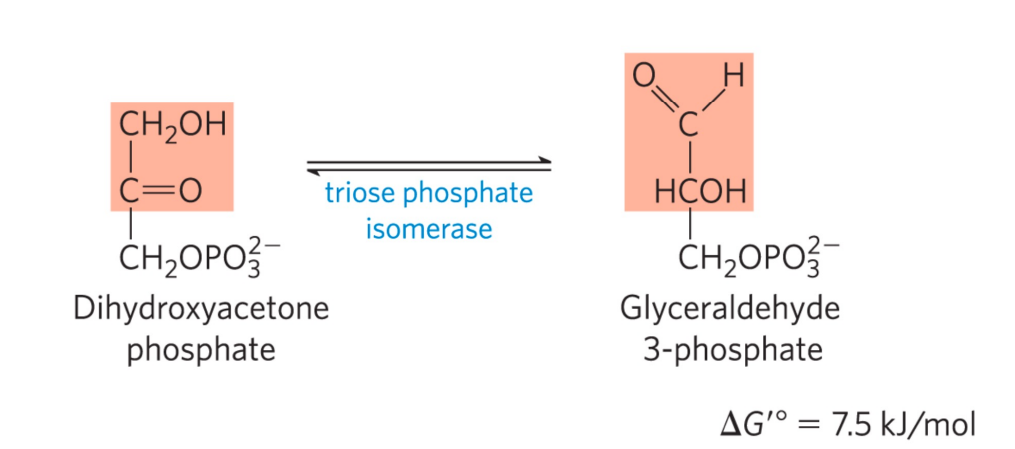
- 本反应是磷酸丙糖异构酶催化的异构反应。
- 属于分子内氧化还原反应。
Hold on...
至此我们走完了糖酵解的前五步准备阶段,一分子葡萄糖总计消耗了两分子的ATP,生成了一分子的磷酸二羟丙酮与一分子的3-磷酸甘油醛。
当然,下一步反应只接受3-磷酸甘油醛,而这两者又有异构反应的存在,所以我们也可以认为准备阶段生成了两分子的3-磷酸甘油醛。
第六步: 氧化3-磷酸甘油醛
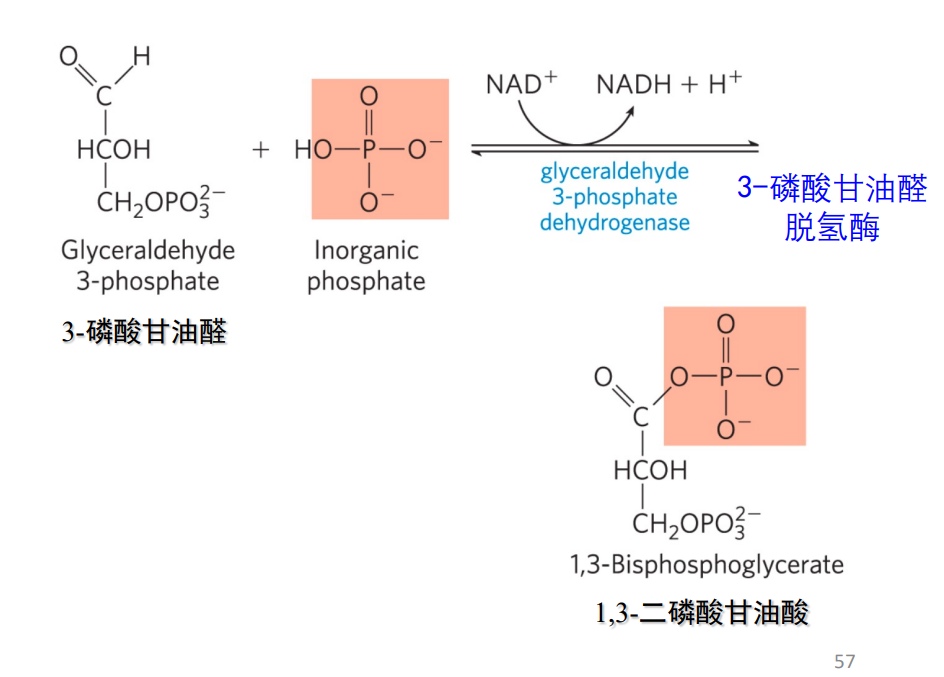
- 该反应由3-磷酸甘油醛脱氢酶催化,3-磷酸甘油醛被氧化,\(\ce{NAD+}\)被还原为\(\ce{NADH}\)。
- 生成的1,3-二磷酸甘油酸是酰基磷酸,具有较高的磷酰基团转移潜能,可以转移给ADP。
Note
辅酶I 是烟酰胺腺嘌呤二核苷酸,具有氧化型与还原型(NAD+/NADH)。
辅酶II 是烟酰胺腺嘌呤二核苷酸磷酸,也具有氧化型与还原型(NADP+/NADPH)
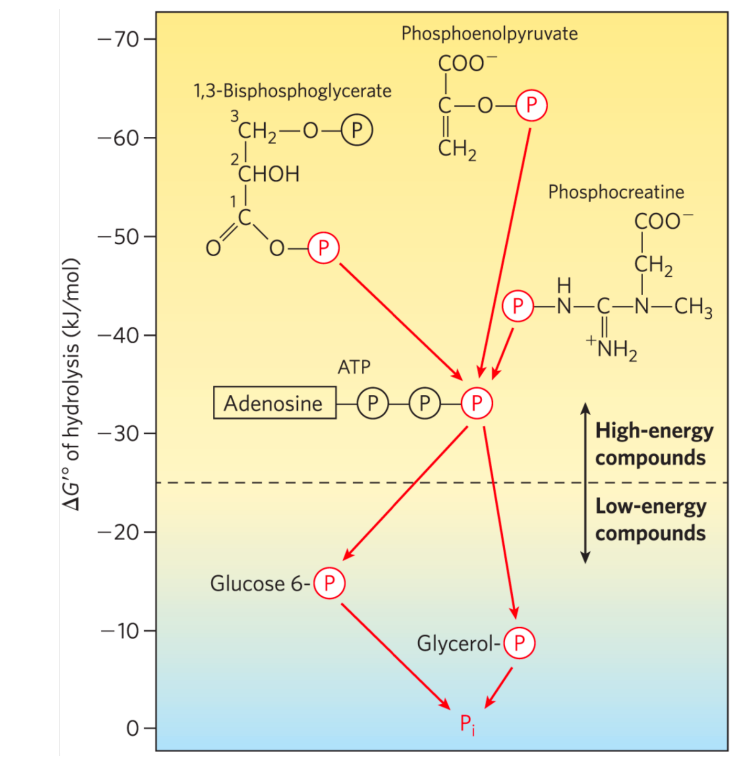
1,3-二磷酸甘油酸的磷酰基团有很高的转移势能,换句话说,它具有高能磷酸键,1,3-二磷酸甘油酸也是生物体内除了ATP以外唯三含有高能磷酸键的小分子物质。
这也意味着由它可以生成ATP。
ATP作为重要的能量货币,其高能磷酸键的能量不大不小,成为重要的衔接高能量,到平稳释放出低能量的物质。
第七步: 磷酸甘油酸激酶催化底物水平磷酸化
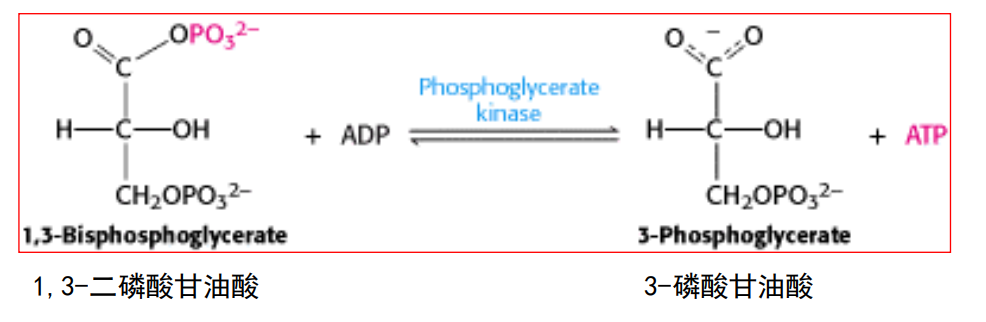
- 这是第一次底物水平磷酸化(与氧化磷酸化区分)
- 催化本反应的磷酸甘油酸激酶是由逆反应命名的。
第八步: 磷酸甘油酸变构酶催化异构
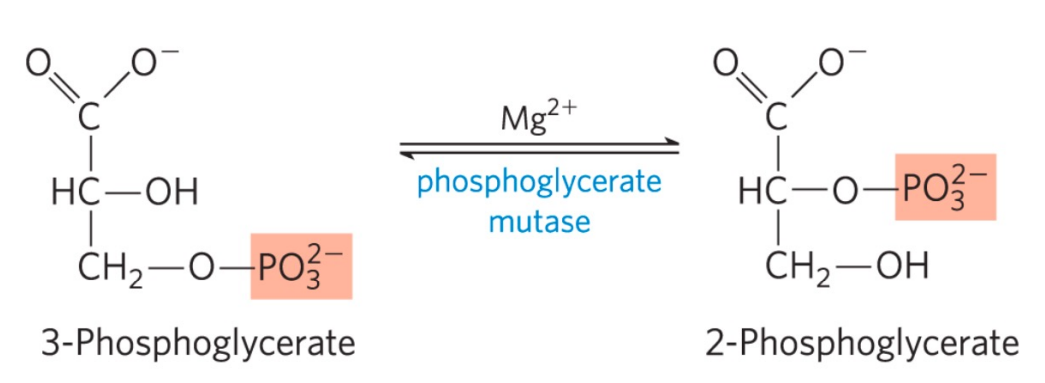
- 该反应由磷酸甘油酸变构酶催化,酶催化分子内基团转移。
第九步: 烯醇化酶催化生成磷酸烯醇式丙酮酸
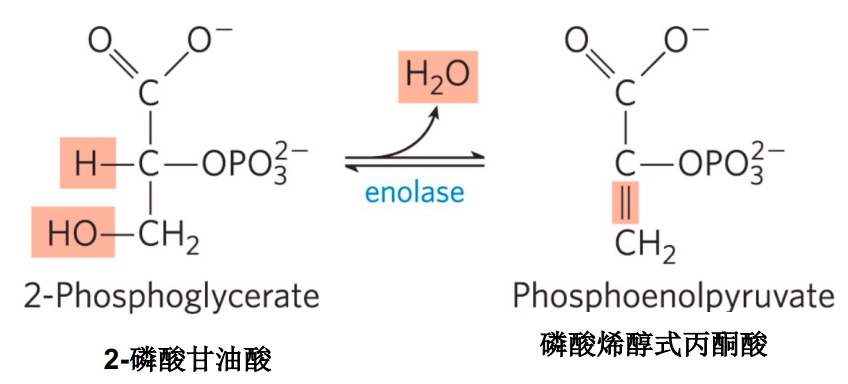
- 本步反应为脱水反应,磷酸烯醇式丙酮酸具有高磷酰基团转移潜能。
第十步: 丙酮酸激酶催化底物水平磷酸化
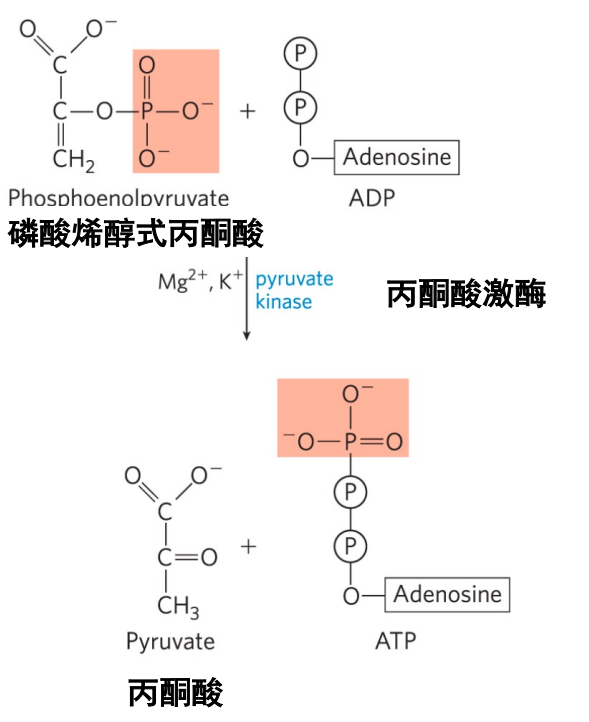
- 本反应为不可逆反应,为第二步底物水平磷酸化。
糖酵解的最后
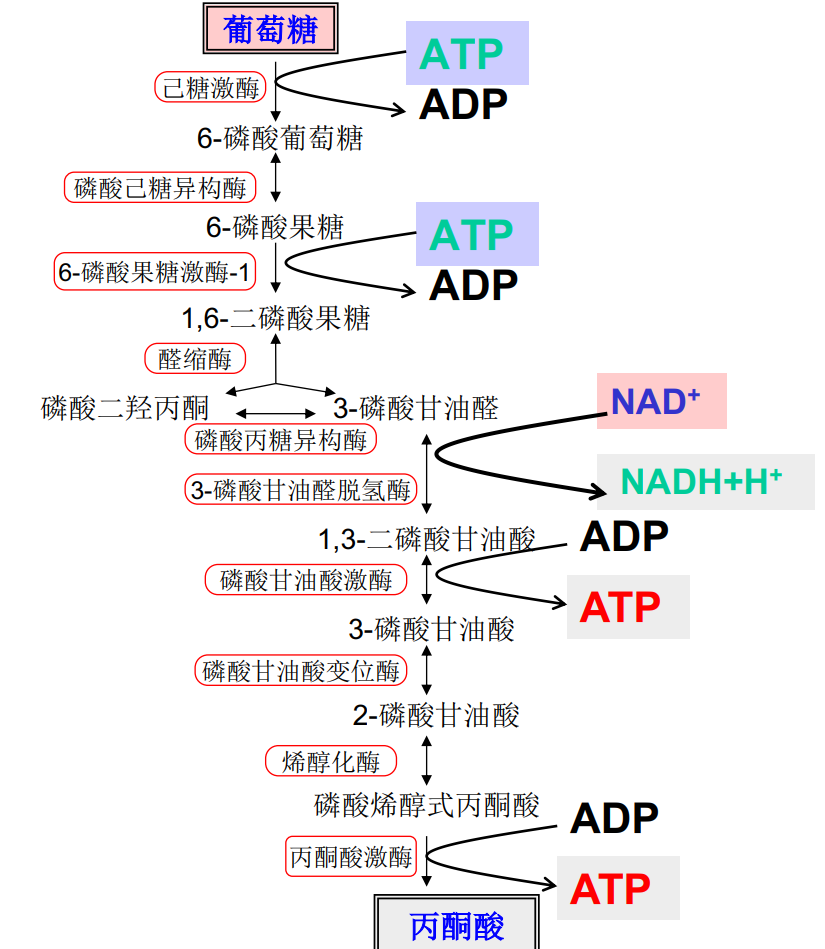
最终,一分子葡萄糖净生成2分子ATP,2分子NADH。
糖酵解要点
- 十步反应,准备阶段五步,收获阶段五步。
- 准备阶段生成磷酸二羟丙酮 (DHAP) 和3-磷酸甘油醛 (GAP)。
- 生成 NADH 时将醛氧化为酸。
- 磷酰基团和磷酸烯醇式丙酮酸携带高能磷酸键。
2,3-二磷酸甘油酸旁路
红细胞中存在2,3-二磷酸甘油酸旁路,从而积累了大量的2,3-二磷酸甘油酸。
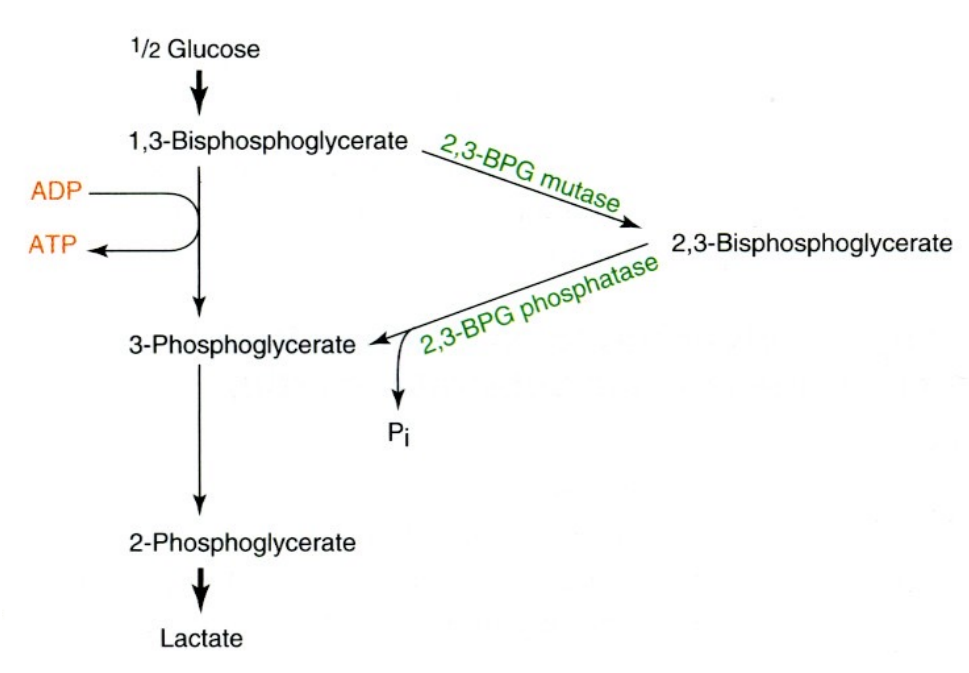
糖酵解的调节位点
- 己糖激酶,PFK-1与丙酮酸激酶是糖酵解的重要调节酶。
- 一般而言,代谢途径中催化不可逆反应的酶是重要的调节位点
- 催化效率低的限速酶往往也是重要的调节位点
酶活性常见方式由快到慢为:变构调节、共价修饰和表达水平。
已糖激酶同工酶
已糖激酶存在四种同工酶,HK I到 HK IV。
<待补充>
丙酮酸的命运
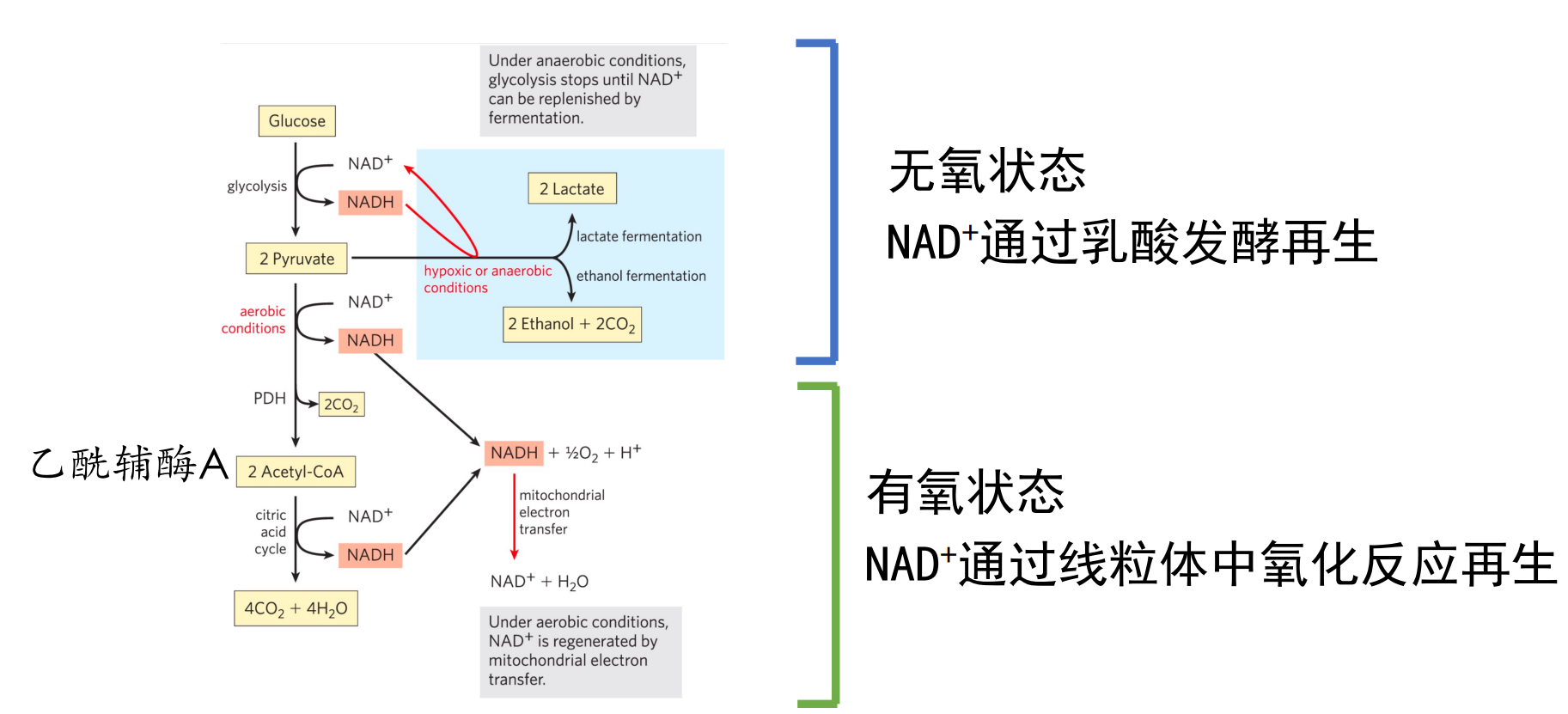
在有氧条件下,丙酮酸会被完全氧化为二氧化碳与水,而在无氧条件下,丙酮酸的命运则取决于生物体本身的倾向。
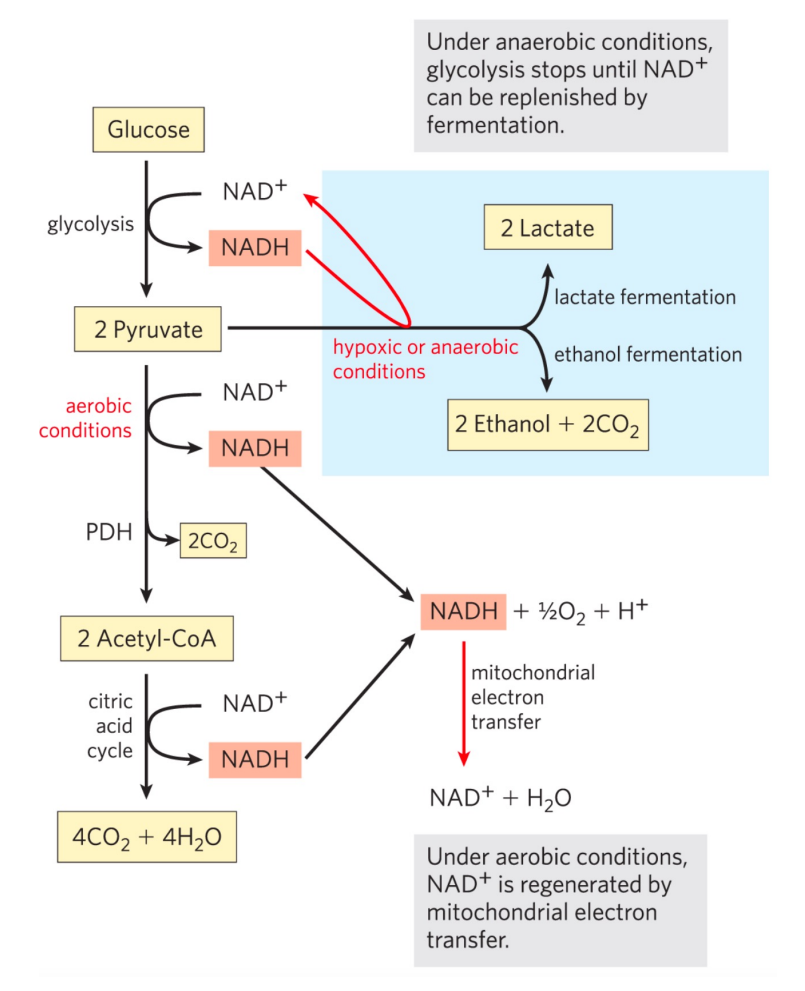
乳酸脱氢酶
高等生物中会通过乳酸脱氢酶将丙酮酸催化还原为乳酸,消耗了糖酵解生成的\(\rm{NADH}\),实现了\(\ce{NAD+}\)的再生。
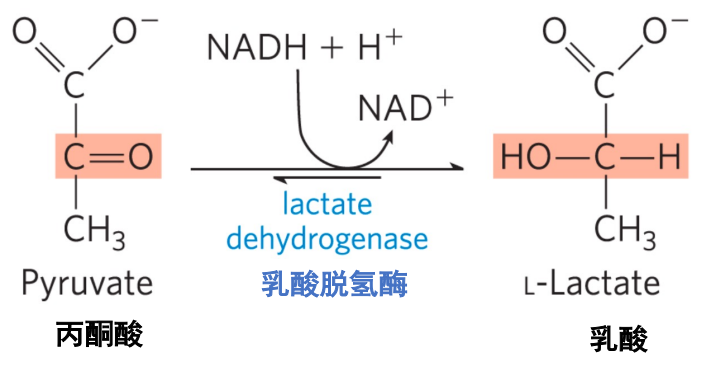
Note
大量文献中也将葡萄糖到乳酸的途径称为糖酵解。
部分厌氧微生物则完全依赖于这种途径来产生ATP,比如制作泡菜用到的乳酸菌。
丙酮酸脱氢酶与乙醇脱氢酶
酿酒酵母等微生物可以利用丙酮酸脱氢酶、乙醇脱氢酶来催化丙酮酸生成乙醇。
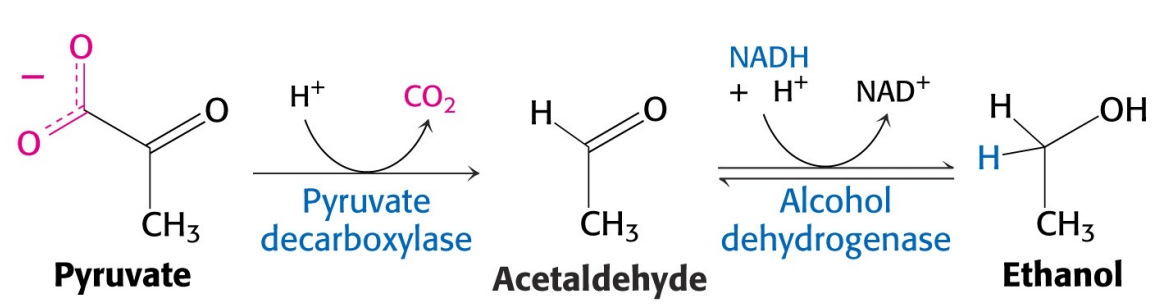
<待补充>
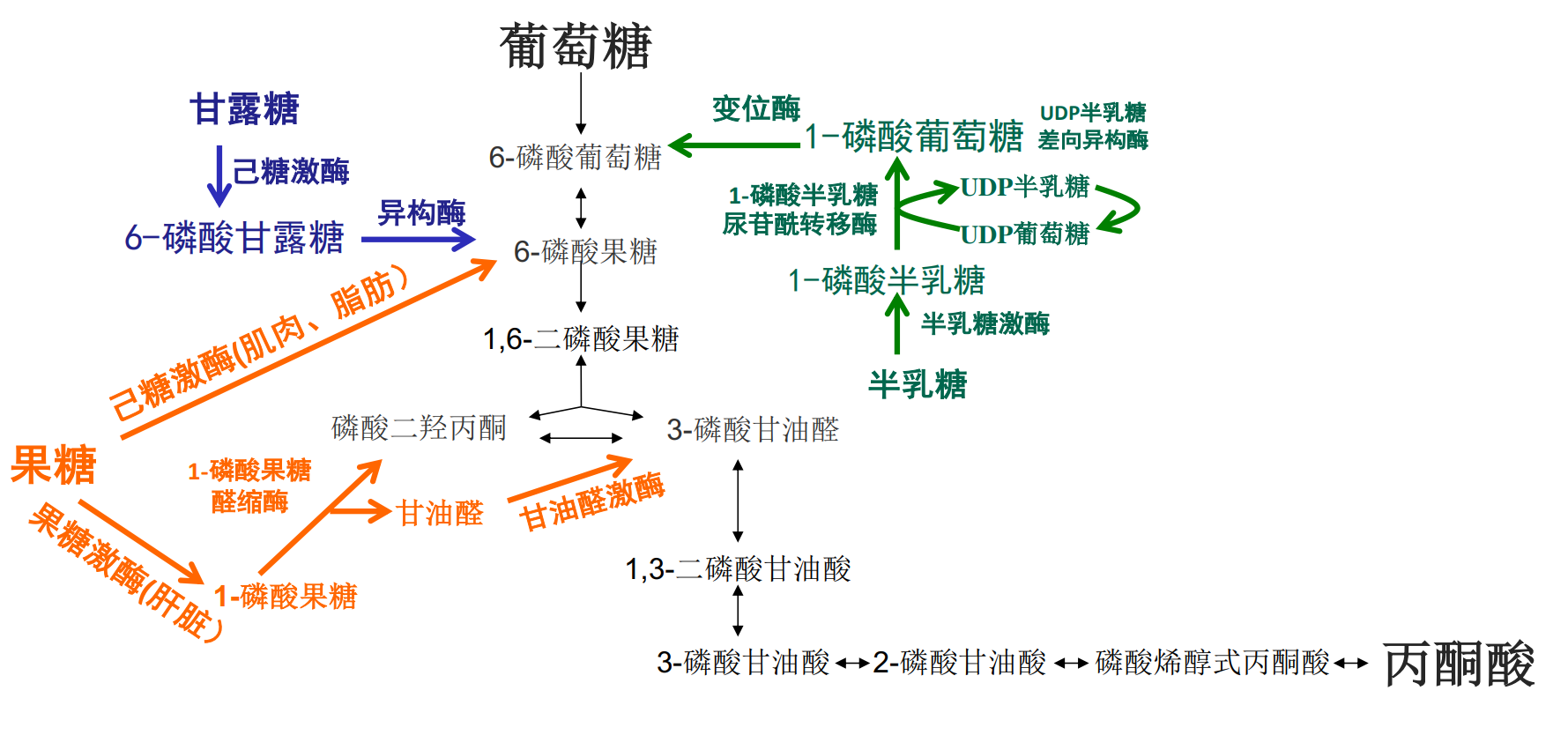
其他单糖进入糖酵解的途径
人体能摄取的单糖可不只有葡萄糖,半乳糖、甘露糖与果糖在体内转变成糖酵解途径的代谢中间体,从而进入糖酵解代谢途径。
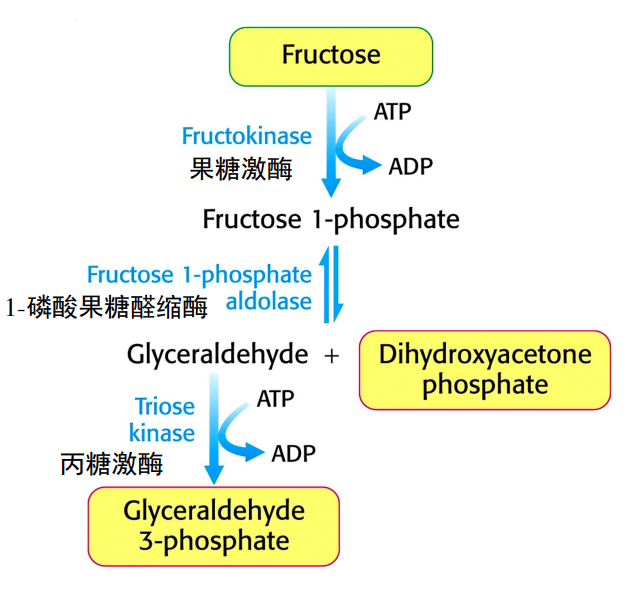
果糖
肝脏是果糖的主要代谢器官,通过果糖激酶催化为果糖-1-磷酸,再由1-磷酸果糖醛缩酶催化生成磷酸二羟丙酮与甘油醛。甘油醛会在丙糖激酶的催化下生成3-磷酸甘油醛,从而将果糖转化为糖酵解的代谢中间体。
而在脂肪等组织中果糖主要是由己糖激酶直接转化为果糖-6-磷酸。
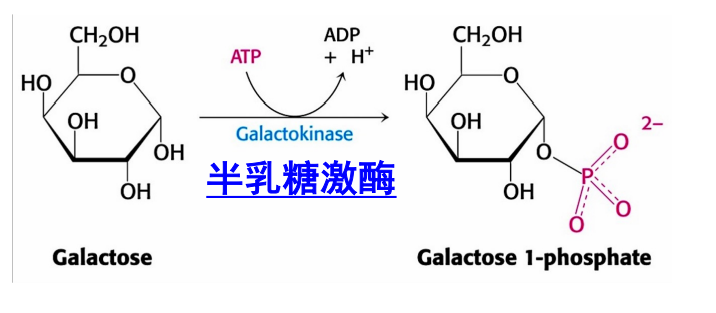
半乳糖
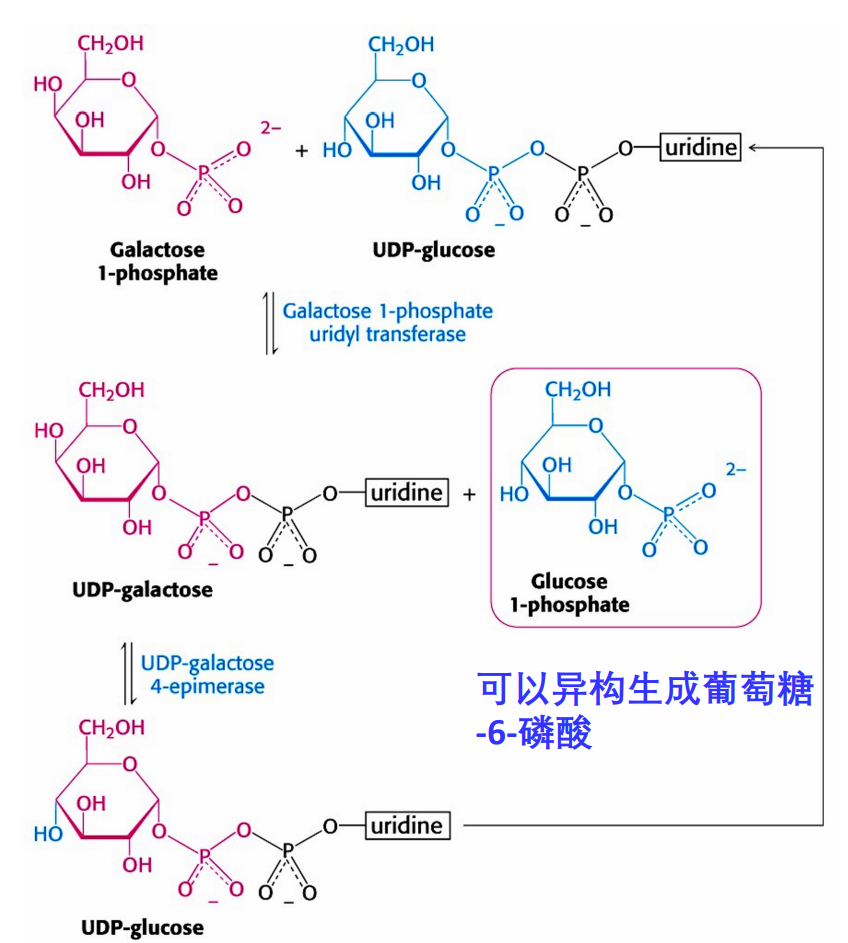
半乳糖首先被半乳糖激酶转化为半乳糖-1-磷酸,再与UDP-葡萄糖在半乳糖-1-磷酸尿苷转移酶(GALT)的催化下生成葡萄糖-1-磷酸与UDP-半乳糖,前者异构为葡萄糖-6-磷酸,后者异构为UDP-葡萄糖。
总结
下图展示了糖酵解在不同单糖中的重要地位。
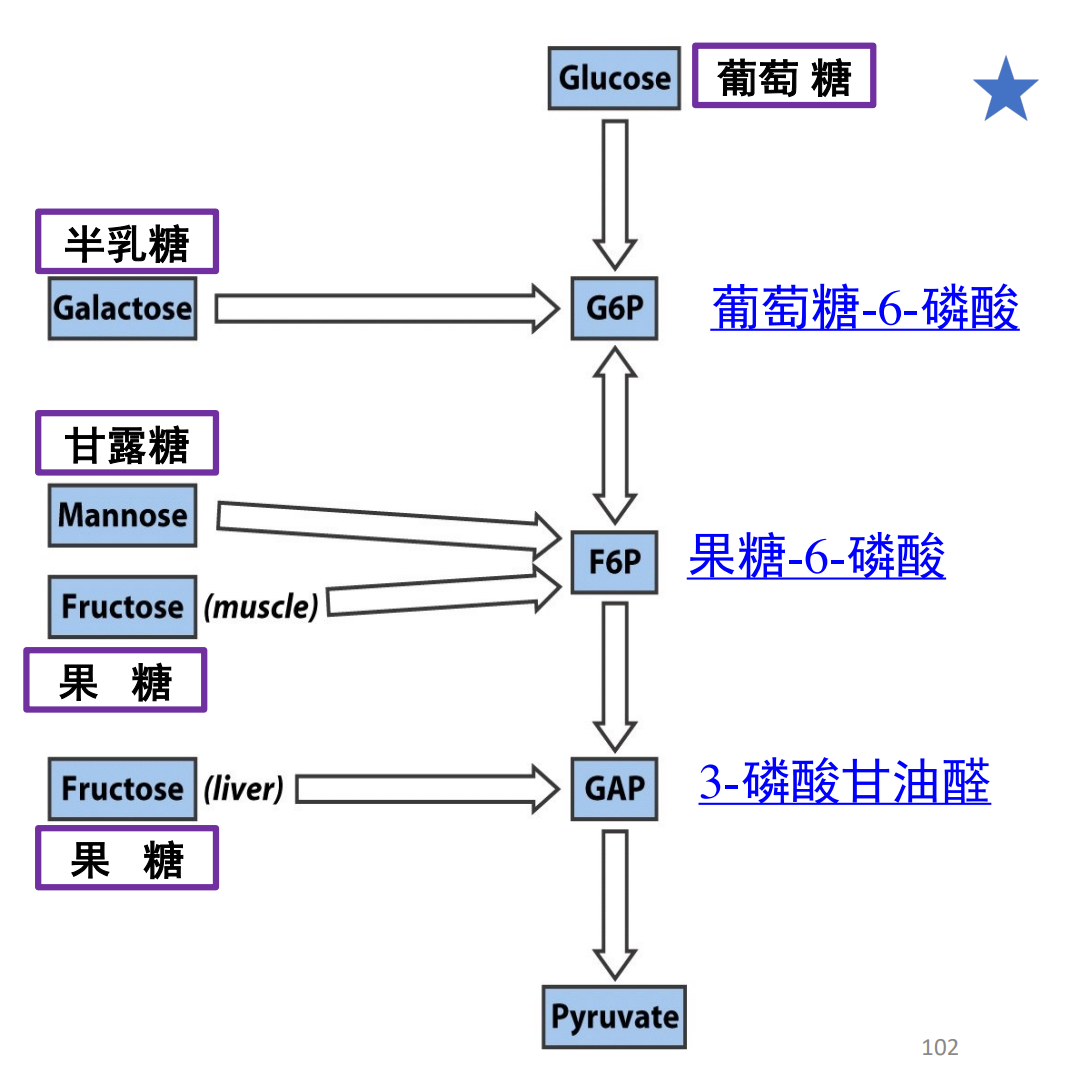
最后可以概括为这张图:
葡萄糖的有氧氧化
在机体供养充足的时候,葡萄糖彻底氧化为二氧化碳与水,并释放出大量能量的过程被称为葡萄糖的有氧氧化,其是人体的主要功能方式。
糖的有氧氧化的反应过程包括糖酵解途径、丙酮酸氧化脱羧、三羧酸循环、电子传递偶联氧化磷酸化,主要在胞浆和线粒体中完成。
这里我们主要涉及糖酵解之后的有氧部分。
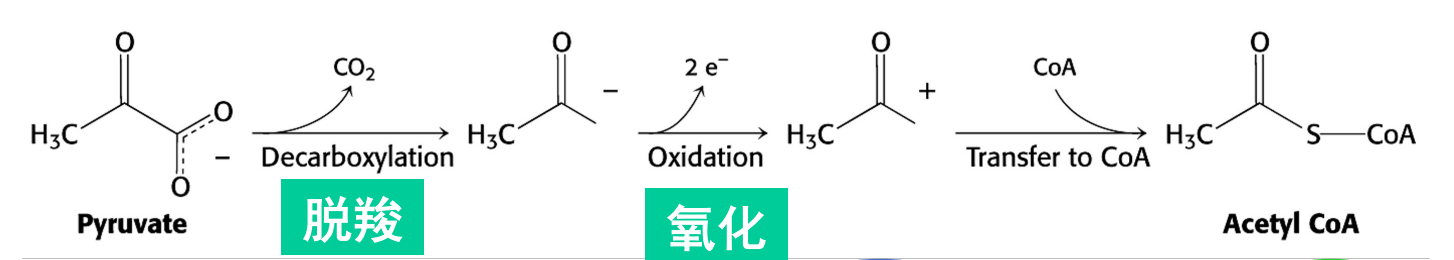
丙酮酸脱羧氧化生成乙酰辅酶A
丙酮酸的脱羧氧化在线粒体基质中进行,其通过丙酮酸转运体进入线粒体。
主导丙酮酸脱羧氧化反应的是丙酮酸脱氢酶复合体,其可以催化丙酮酸转化为乙酰辅酶A,并生成1分子二氧化碳与NADH。
丙酮酸脱氢酶复合体
丙酮酸脱氢酶复合体包含三种酶单体:
- 丙酮酸脱氢酶(E1)与辅酶TPP
- 二氢硫辛酰胺转乙酰酶(E2)与辅基硫辛酸
- 二氢硫辛酰胺脱氢酶(E3)与辅酶NAD,FAD
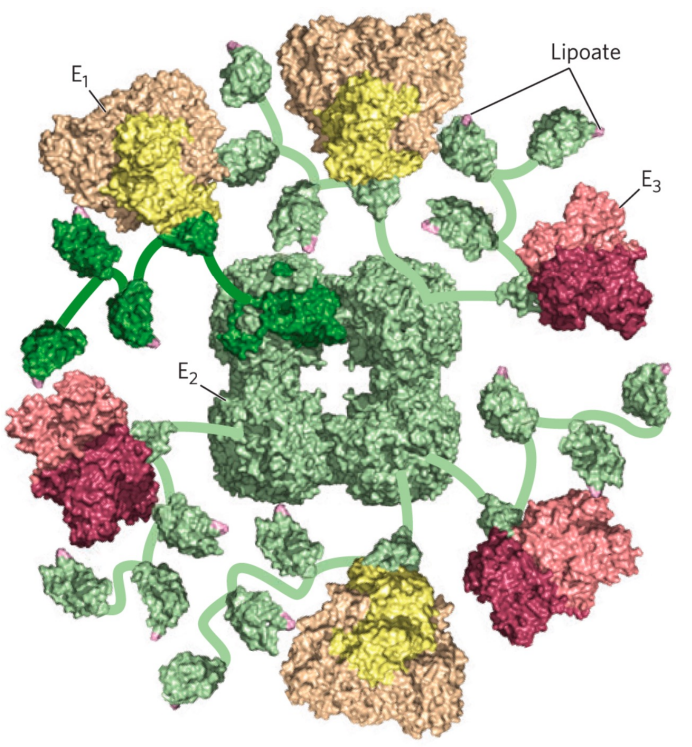
其过程必需五种辅酶的参与:NAD(烟酸,维生素B3,作为氢和电子的受体)、硫胺素焦磷酸(硫胺素,维生素B1,TPP)、硫辛酸(lipoate)、FAD(核黄素,维生素B2)、辅酶A(泛酸,维生素B5,CoA)。
糖异生
葡萄糖是部分哺乳动物的唯一或主要功能分子,例如人的脑组织与神经系统、睾丸、肾髓质、红细胞和胚胎组织在正常条件下只使用葡萄糖。鉴于葡萄糖在人体内的重要地位,葡萄糖在体内的缺乏是难以被接受的。
幸好生物体内存在糖异生这一途径——非糖物质转变葡萄糖或糖原的过程。在人体内主要由肝脏进行糖异生,肾皮质与小肠上皮细胞也可以进行糖异生,但成熟红细胞与脑不能进行。
Note
- 糖异生不是糖酵解途径的逆过程。后者有三步不可逆反应。
- 从丙酮酸生成葡萄糖,有四步独特的反应与酶,其他与糖酵解共用。糖异生通过这四步反应来跳过糖酵解的不可逆反应。
- 全程在细胞液、线粒体和内质网三个部位完成。
丙酮酸转化为磷酸烯醇式丙酮酸
这一步是糖异生的第一步,需要分为两个阶段进行。
第一步:在线粒体中的丙酮酸羧化酶以丙酮酸为底物催化生成草酰乙酸。
丙酮酸羧化酶
丙酮酸羧化酶与生物素共价结合催化该反应,其是糖异生反应的关键调节酶,受到CoA的正向调节。
第二步:磷酸烯醇式丙酮酸羧激酶在细胞质中催化草酰乙酸转化为磷酸烯醇式丙酮酸的反应。
狸猫换太子
丙酮酸一开始在线粒体基质中,其被原地转化为草酰乙酸后需要转移至细胞质基质进行下一阶段的反应。
但是问题来了,线粒体的内膜对草酰乙酸不具通透性——内膜通透性低,且不存在草酰乙酸的转运体。
线粒体对此的解法是——苹果酸-天冬氨酸穿梭系统。
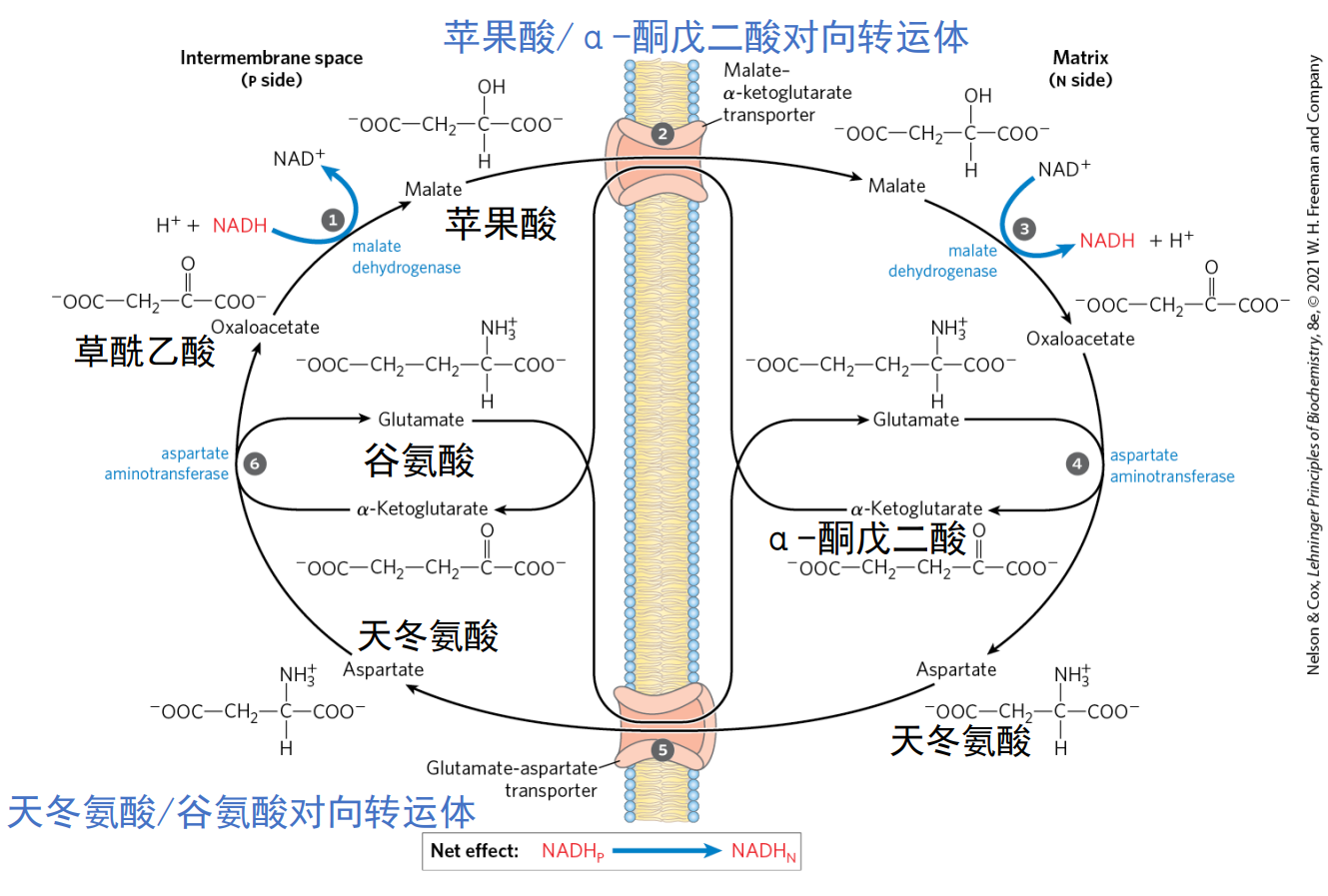
- 草酰乙酸首先通过三羧酸循环最后一步的逆反应转变为苹果酸。
- 苹果酸与α-酮戊二酸对向转运,苹果酸离开线粒体。
- α-酮戊二酸与
<待补充>
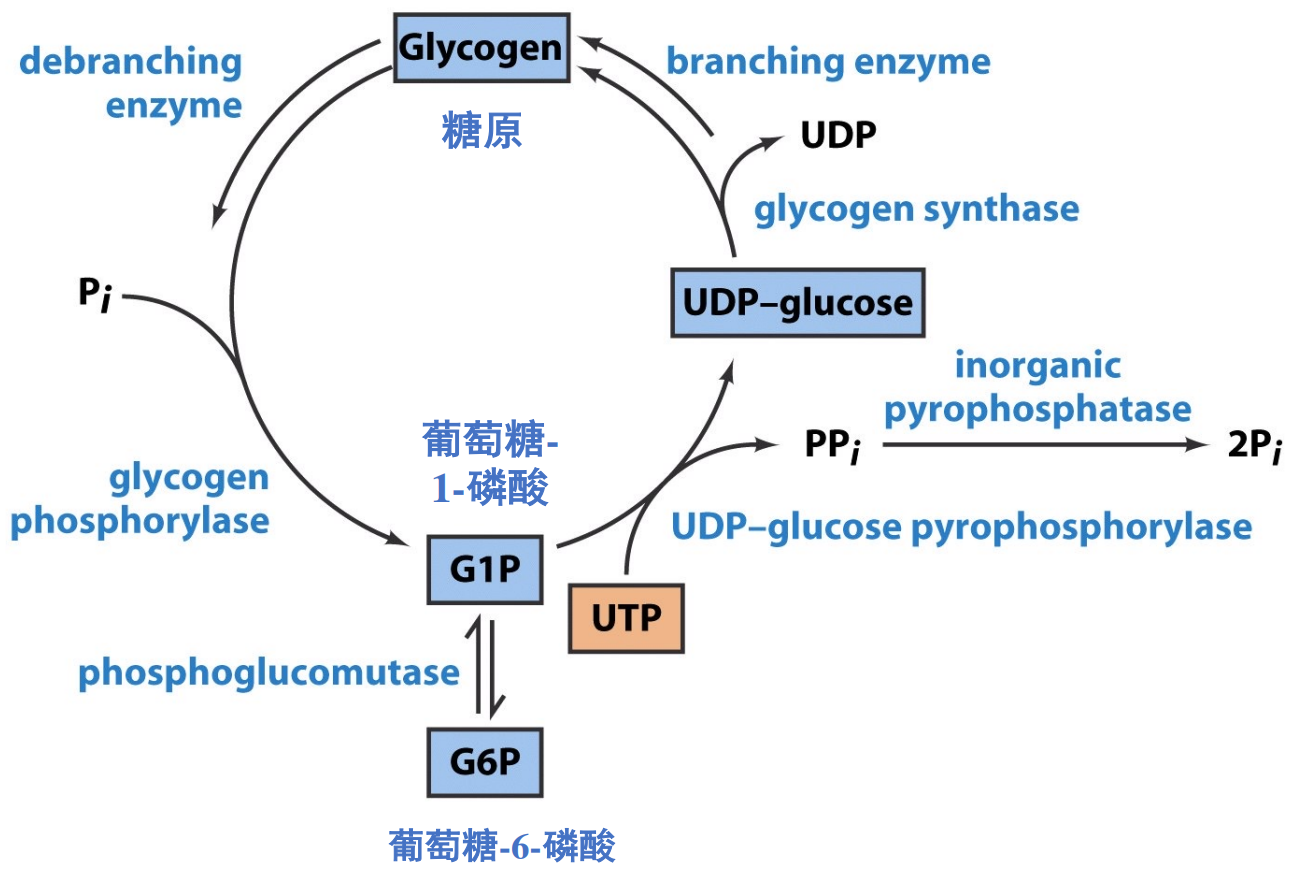
糖原的分解与合成
糖原 (glycogen) 是一种容易动员的葡萄糖储存形式,能在储存的同时不影响细胞的渗透压,是血糖的重要来源。
人体内糖原主要存在于肝脏与肌肉中,分别是肝糖原与肌糖原。
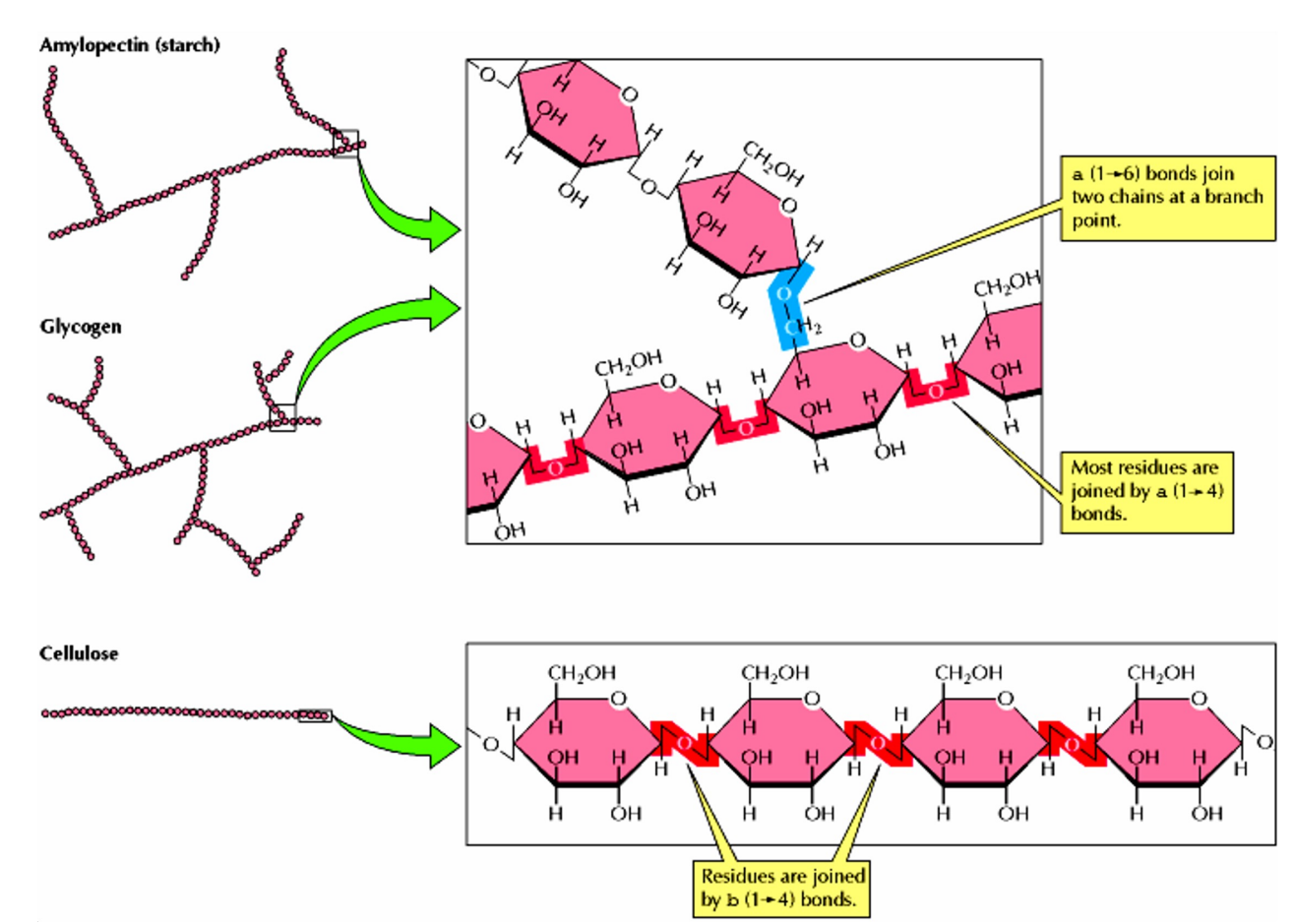
糖原是大的、有分支结构的葡萄糖多聚体,糖原约85%的葡萄糖以α-1,4-糖苷键形成直链,α-1,6-糖苷键形成支链。
糖原的合成
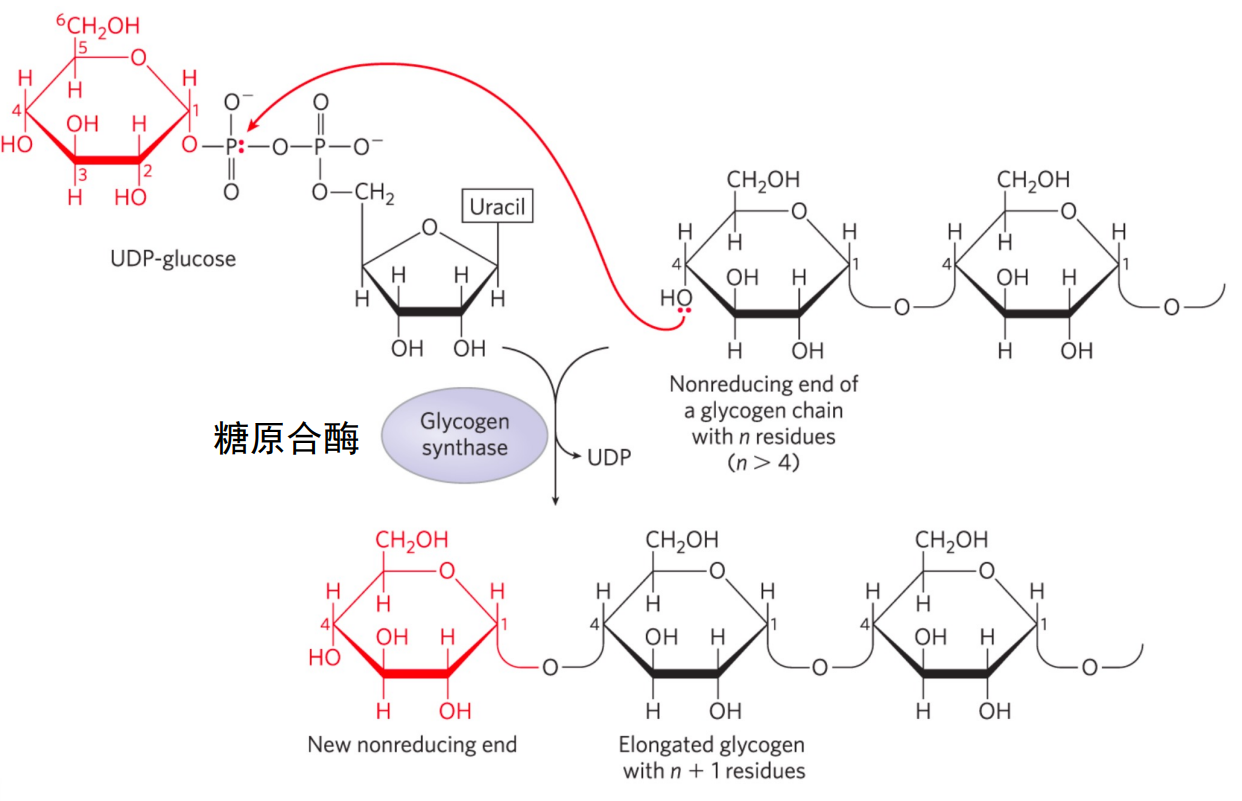
葡萄糖首先要被激活为UDP-葡萄糖后才能参与糖原合成。
- 葡萄糖经糖酵解第一步后生成葡萄糖-6-磷酸,后可变位为葡萄糖-1-磷酸
- UTP与G-1-P在UDPG焦磷酸化酶作用下生成UDP葡萄糖。
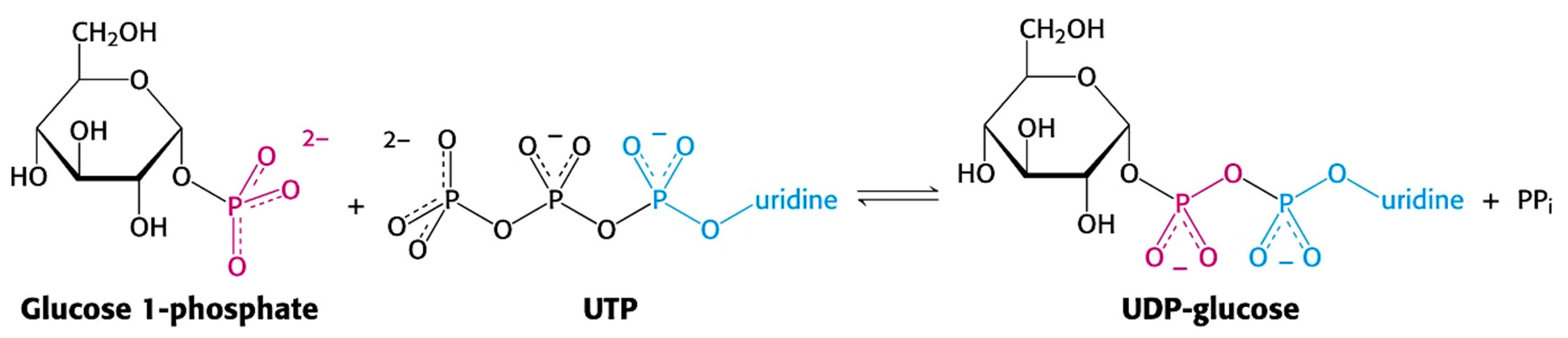
UDP-葡萄糖为何“堪当大任”?
以UDP-葡萄糖作为糖原合成的底物具有如下优点:
- UDP-葡萄糖合成所生成的PPi水解使其反应不可逆,推动合成代谢的正向进行。
- 核苷酸部分促进与酶的非共价结合,同时也是很好的离去基团,促进亲核进攻的发生。
- 与糖酵解反应所需葡萄糖区分开来,“专款专用”
糖原合酶催化UDP-葡萄糖形成α-1,4-糖苷键,延长糖原。
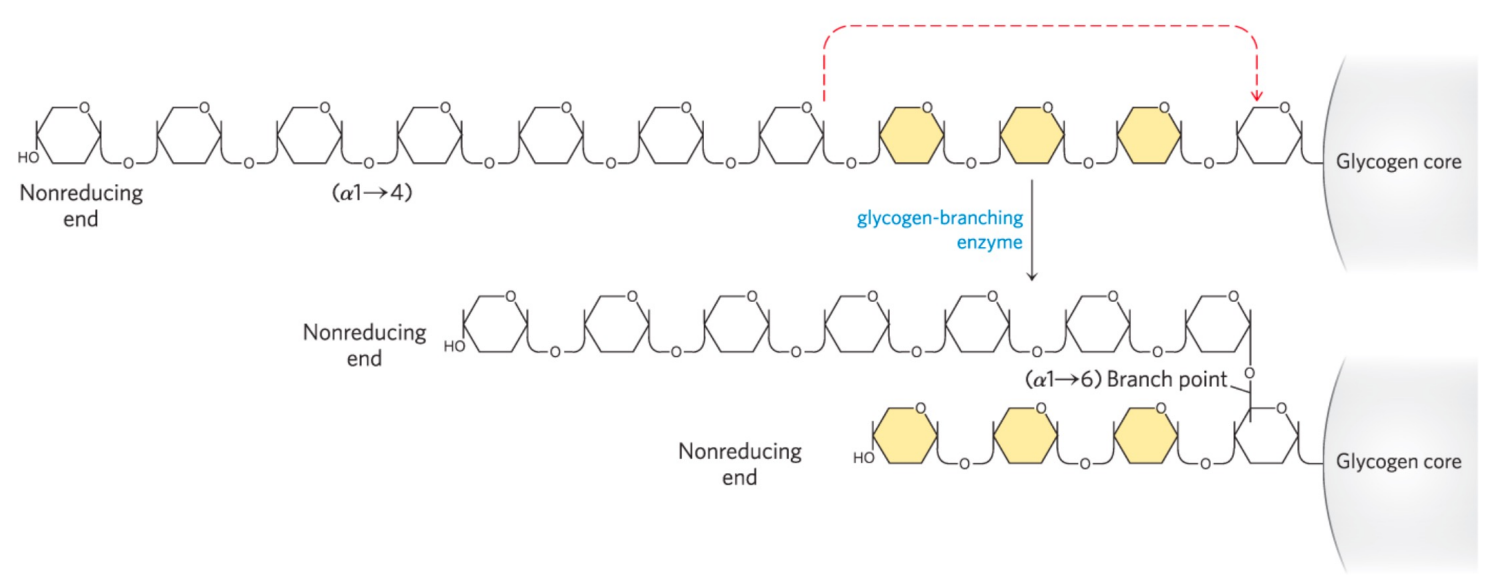
当糖链长度超过11时,分支酶 (branching enzyme),将末端的6-7个葡萄糖残基转移到邻近的糖链上,形成分支。
形成分支的重要生物学意义在于:增加了非还原末端的数量,增加了糖原合成和分解的效率。
糖原的合成原料除了直接从葡萄糖合成,还有一个更为常见三碳途径——大部分葡萄糖先在肝外被转化为乳酸或丙酮酸等三碳化合物,再进入肝细胞异生为糖原的过程。
糖原核心结构
特别的是,糖原合酶不能催化糖原链的从头合成,需要从一个糖原引物开始合成。
糖原生成蛋白 (glycogenin) 通过催化自身残基修饰同时扮演了核心和引物的功能。
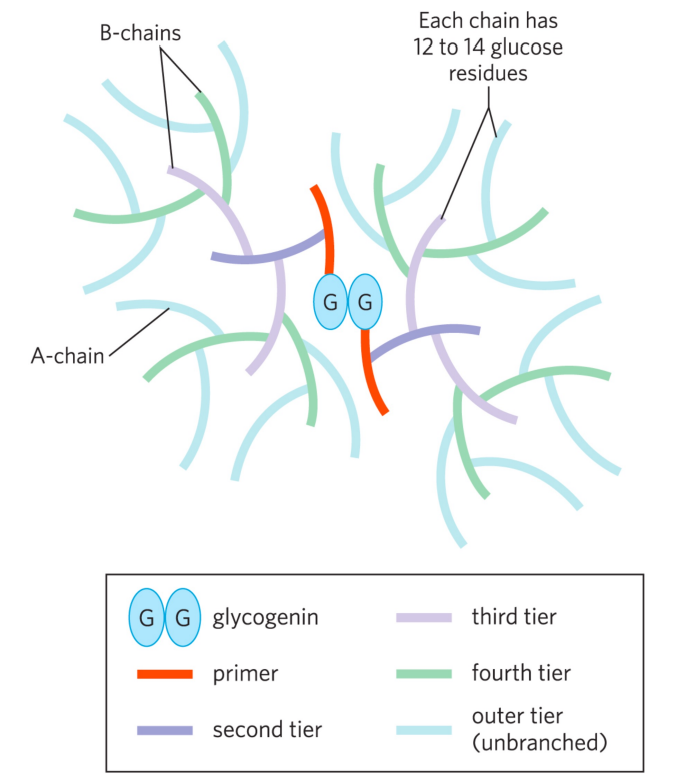
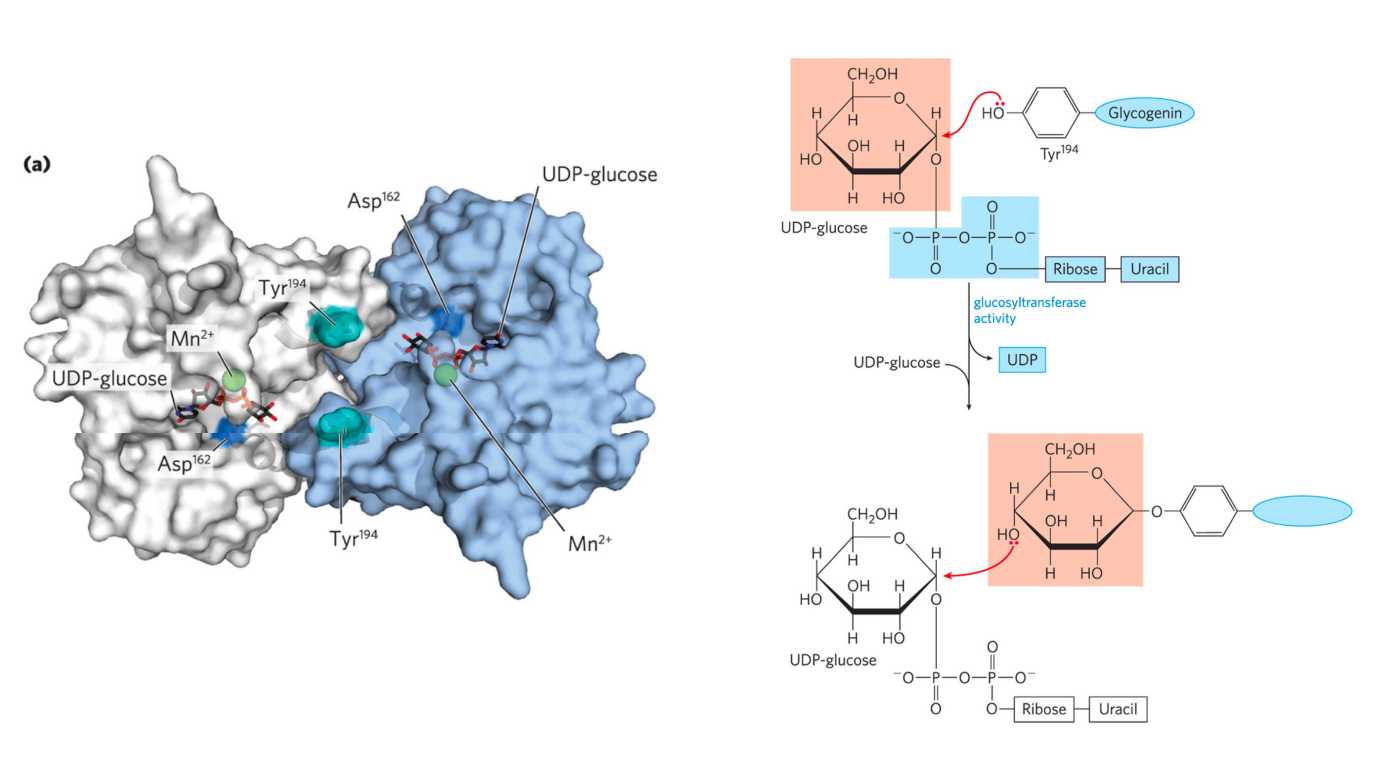
糖原的分解
糖原磷酸化酶催化糖原磷酸解,释放出葡萄糖-1-磷酸。
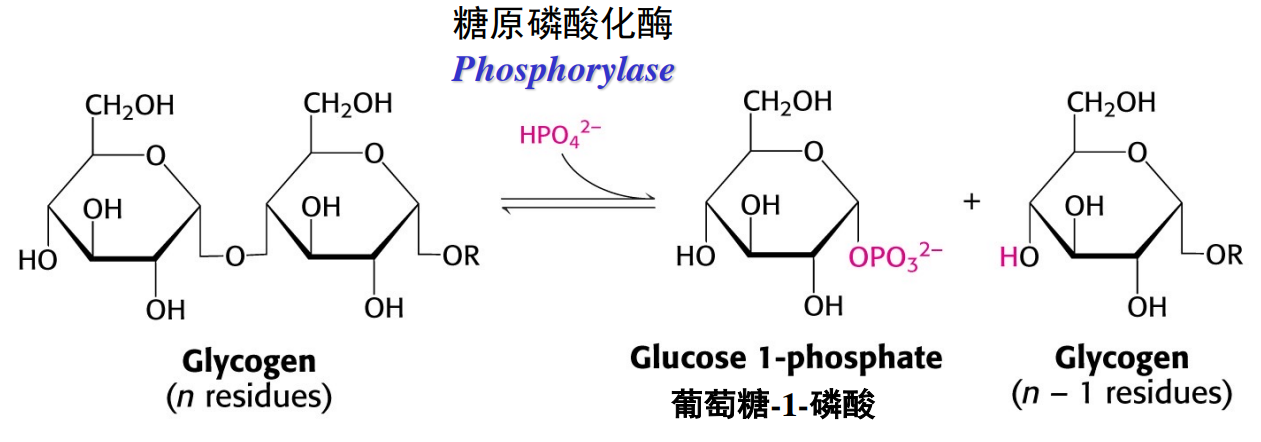
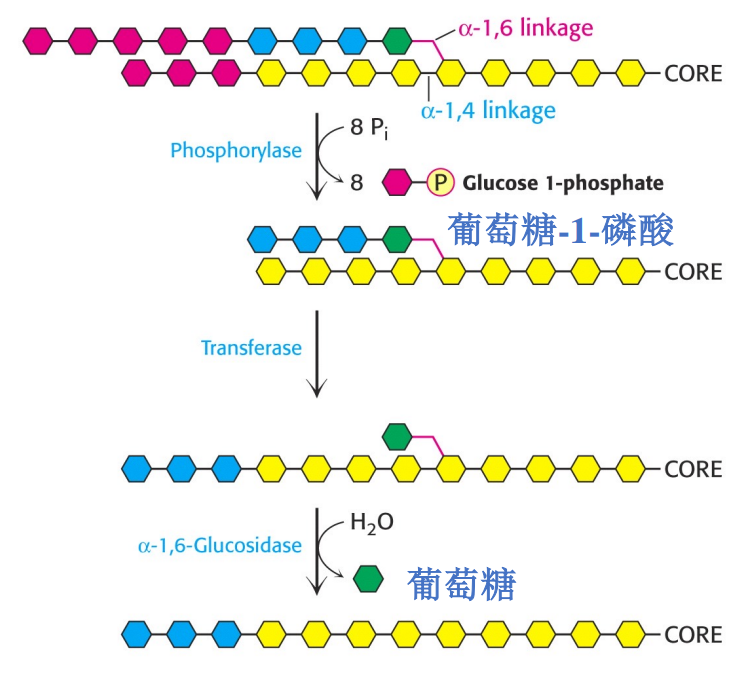
糖原支链则由脱支酶催化分解,其同时具有葡聚糖转移酶和α-1,6-葡萄糖糖苷酶两种活性。
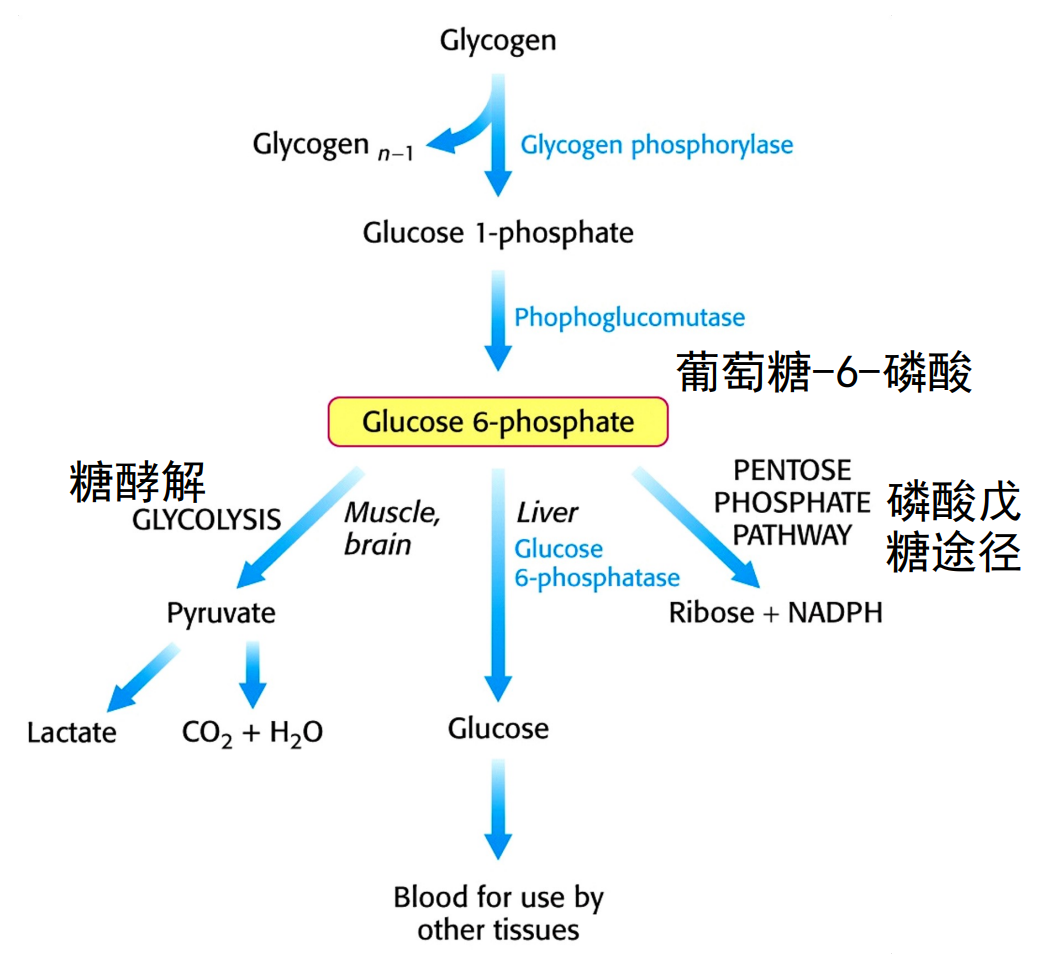
糖原分解形成的葡萄糖-1-磷酸会被变位为葡萄糖-6-磷酸,之后会有三个不同的命运:糖酵解、磷酸戊糖途径与转变为葡萄糖。
Note
只有肝脏才会表达葡萄糖-6-磷酸酶,也就只有肝糖原的分解才能影响到血糖。肌肉中糖原不具有转变为葡萄糖的条件。
糖原合成与分解的调节
糖原分解的关键酶糖原磷酸化酶与糖原合成的关键酶糖原合酶是关键的调控位点,两者在调节上是相反的,可逆磷酸化、激素和变构效应剂是调控的主要机制。
总结
糖原的分支多,除了中心所包含的一个还原端外,存在有众多非还原端,糖原的合成与分解均从非还原端开始。