细胞膜及其表面分子
掌握
- 细胞膜的组成和结构特征。
- 蛋白质与膜结合的方式和膜蛋白的基本特性。
了解
- 膜脂成分的基本生化特性。
- 细胞膜上的特异结构及功能。
细胞质膜及其表面结构:质膜(plasma membrane)包在细胞外面,所以又称细胞膜(cell membrane),它不仅是区分细胞内部与周围环境的动态屏障,更是细胞物质交换和信息传递的通道。 围绕各种细胞器的膜,称为细胞内膜。 质膜和内膜在起源、结构和化学组成的等方面具有相似性,故总称为生物膜(biomembrane)。
生物膜是细胞进行生命活动的重要物质基础。
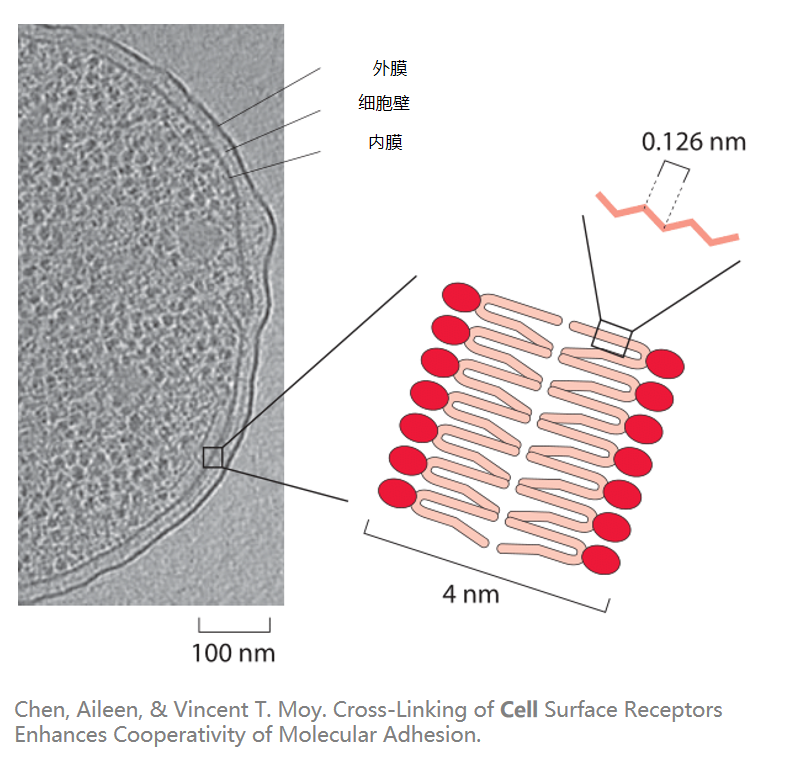
下图展示了一个支原体细胞的微观结构。

生物膜
生物膜(Biological membrane)是对生物体内所有膜结构的统称。它是一层封闭的、有分隔作用的膜,在生物体中担任选择透过性屏障。
细胞膜是生物膜的一种,通常由磷脂双分子层组成,其厚度为4~10nm之间。
生物膜——生命的起源与进化中最重大事件之一
生物膜将“特殊物质集团”区别于周围环境,同时膜又可以吸收周围的营养物质,并将膜内的废物排出去,如此演化出具生命力的细胞。膜是生命最基础的结构。
对生物膜的认知由早期简单的脂类膜到最后的流动镶嵌模型,确认了生物膜有以下性质:
- 膜结构具有流动性
- 膜组分分布不对称
- 膜蛋白以不同的镶嵌方式与膜结合
流动镶嵌模型认为磷脂双分子层构成膜的主体,球状蛋白镶嵌或者附着在脂双层中间及表面,但这仍有不足,因为其忽视了膜流动性的不均一性与膜蛋白对膜流动性的影响。
脂筏模型
脂筏模型是对流动镶嵌模型的补充与发展,脂筏是质膜上富含胆固醇和鞘磷脂的微结构域,其中聚集一些特定种类的蛋白质。其是一种动态结构,位于质膜磷脂双分子层的外侧,是细胞膜中的功能化筏。
脂筏与小窝蛋白(caveloin)结合后内陷形成小窝,参与跨膜物质转运,同时与膜的信号转导、蛋白质分选均有密切的关系。
脂筏具有如下特点:
- 许多蛋白质聚集在脂筏内,便于相互作用
- 脂筏提供一个有利于蛋白质变构的环境,使形成有效的构象。
生物膜的基本结构特征
生物膜有如下特征:
- 生物膜是所有细胞的共同特征
- 膜主要是由脂类(lipid) 和蛋白质以非共价键相互作用结合而成的二维流动体系。
- 脂类分子呈连续的双分子层(bilayer)排列。
- 膜具有双亲性。
- 蛋白质相对于脂双层具有不同镶嵌方式。
- 生物膜中各种组分的分布是高度不对称的。
膜的不对称性
膜具有不对成性,分别是膜蛋白分布的不对称、膜脂分布的不对称和膜糖分布的不对称,他们决定的是膜功能的方向性。
膜脂内外分布具有不对称性,由于脂分子不能自发地在单层与单层之间以适当的速度翻转,这种不对称可能在内质网合成时已经开始了。脂双层的不对称性有重要的生物学功能!
- 外层 - glycolipids - phosphatidylcholine(PC 磷脂酰胆碱) - cholesterol
- 内层 - phosphatidylserine (PS 磷脂酰丝氨酸) - phosphatidyl-ethanolamine (PE 磷酯酰乙醇胺) - phosphatidylinositol (PI 磷脂酰肌醇)
膜在侧向分布上也具有不均匀的性质。
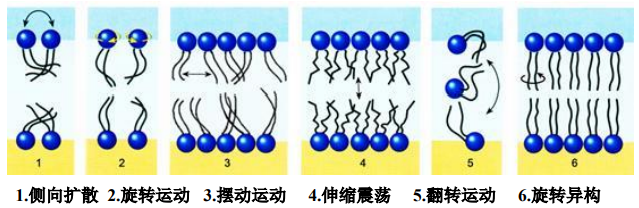
膜的流动性
膜上脂分子有以下运动形式:
- 侧向扩散:同一平面上相邻的脂分子交换位置。
- 旋转运动:膜脂分子围绕与膜平面垂直的轴进行快速旋转。
- 摆动运动:膜脂分子围绕与膜平面垂直的轴进行左右摆动。
- 伸缩震荡:脂肪酸链沿着与纵轴进行伸缩震荡运动。
- 翻转运动:膜脂分子从脂双层的一层翻转到另一层。是在翻转酶(flippase)的催化下完成。
- 旋转异构:脂肪酸链围绕C-C键旋转,导致异构化运动。
生物膜的主要功能
生物膜将细胞分割为“小室”、具有选择通过性、是“小室”间传递化学信息和能量的介面、为蛋白质的合成、加工与修饰、分选与定位,提供了工作平台和输运载体。
区隔化/房室化
内膜结构将细胞分隔成若干独立空间。在每一个膜包裹的空间聚集着特定种类的生物大分子,共同完成着某一项特定的生命活动。
- 细胞核主要进行DNA的复制和转录。
- 线粒体主要进行呼吸作用,提供能量。
- 溶酶体则负责将一些吸收或损伤的大分子降解为可以利用的小分子等。
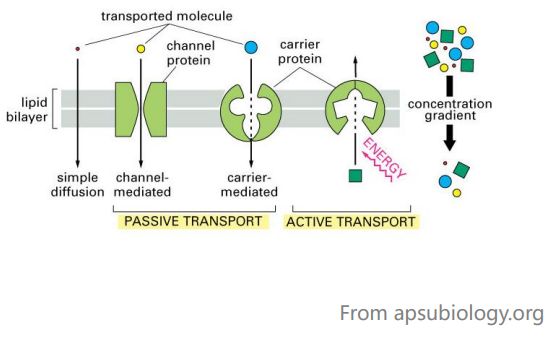
物质的跨膜运输
生物膜既要防止细胞与环境之间以及细胞内各房室之间的物质自由混合,又要维持各区间物质有控制地交流。
能量转换
参与光合作用与呼吸作用。
细胞信息传递
细胞的信息传递指的是细胞通过胞膜或胞内受体感受信息分子的刺激,经细胞内信号转导系统转换,从而影响细胞生物学功能的过程。
- 水溶性信息分子必须首先与胞膜受体结合,启动细胞内信号转导的级联反应,将细胞外的信号跨膜转导至胞内。
- 脂溶性信息分子可进入胞内,与胞浆或核内受体结合,通过改变靶基因的转录活性,诱发细胞特定的应答反应。
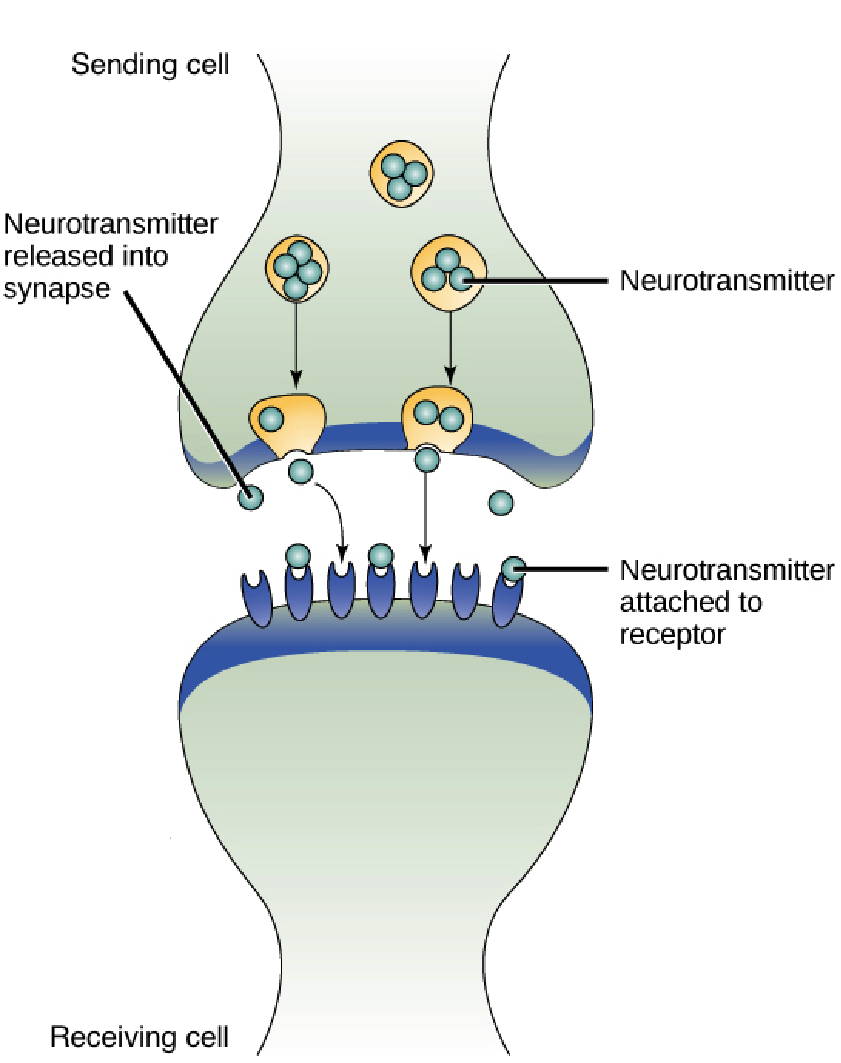
细胞识别
细胞通过其表面的特殊受体与胞外信号物质分子或配体选择性地相互作用,触发细胞内一系列生理生化变化,最终导致细胞的总体生物学效应相应改变,这样的过程称为细胞识别。
生物膜表面分子
生物膜的三种基本组分是:脂类【膜脂(membrane lipid),包含有磷脂、糖脂与固醇,主要是磷脂】、蛋白质(包括酶)和少量糖类组成,也包括水、无机盐、金属离子等。
所有生物膜都具有双亲性,即两侧亲水表面中间夹着疏水的核心。生物膜的所有组分有规则地排列以维持这种双亲结构。
膜脂
细胞中含有两种脂类:
- 非极性脂质:如由脂肪酸与甘油酯化所形成的三酯。它们是一类疏水的脂质,称为真脂。
- 极性脂质:这种脂质分子具有一个亲水的头部(即极性端)和一个疏水的尾部(即非极性端),具有双亲性(amphipathic或amphiphilic)的特点,这种脂质称为类脂。
生物膜中的脂质大多都是极性脂质,包括磷脂(phosnholipid)、固醇(sterol)和糖脂(glycolipid)。
不同细胞器膜的脂类与磷脂组成均不同。
磷脂及磷脂代谢
含有磷酸的脂类统称为磷脂,主要分为两类:
- 甘油磷脂:由甘油构成的磷脂(体内含量最多)
- 鞘磷脂:由鞘氨醇构成的磷脂
两者的分子结构模型示意如下:
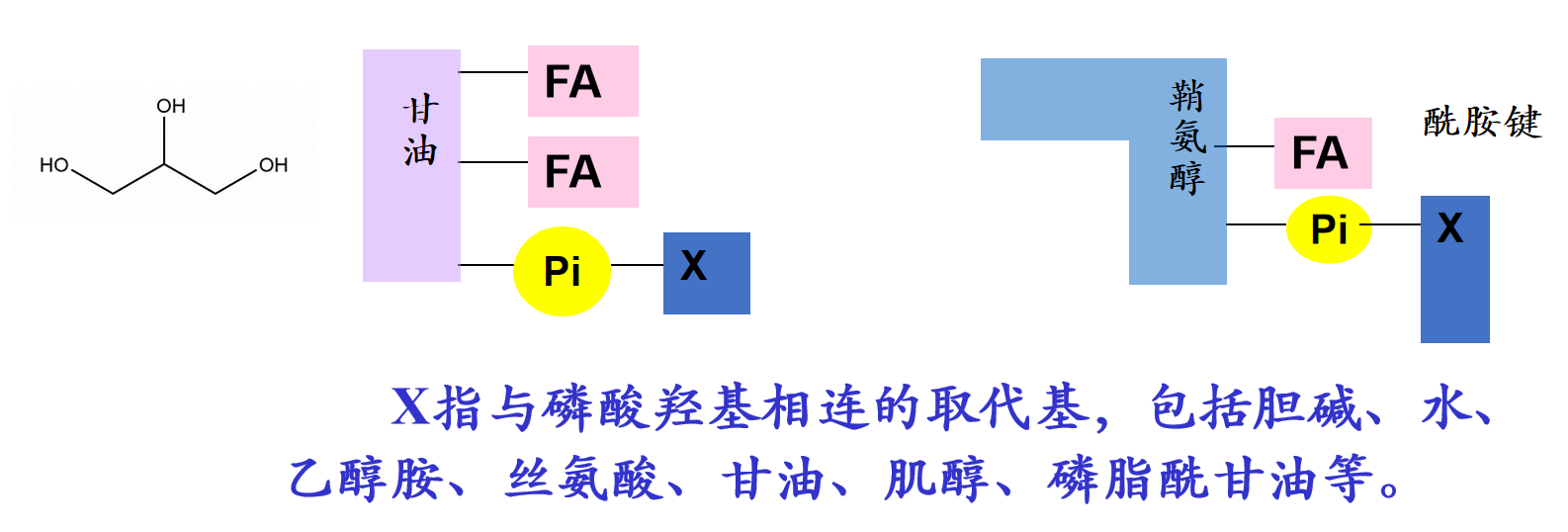
磷脂中含有脂肪酰链,其中可能含有双键(多为顺式构型),含有顺式双键的磷脂在膜上的覆盖面积更大。
磷脂的极性头由带负电荷的磷酸基与带电或不带电的极性基团组成,其决定整个磷脂的电荷特性。而在生理状态下,磷脂在膜内外分布的不均衡使膜内侧带有额外的负电荷。
膜脂不对称性
细胞为什么要维持“膜脂不对称性”?要知道为了形成、维持与调节膜脂不对称性是要消耗宝贵的能量的,细胞要这么做一定有它的原因。
经过一番检索我认为膜脂不对称性,或者说膜内侧为什么要维持为负电荷至少有以下几种作用:
- 参与信号传导,招募信号蛋白与作为信号蛋白的辅助因子。
- 参与细胞凋亡,一般情况下由翻转酶Flippase将带负电荷的磷脂酰丝氨酸控制在细胞膜内侧,但是当细胞凋亡发生的时候,扰乱酶Scramblase出手将磷脂酰丝氨酸翻转至细胞膜外侧,从而暴露出负电荷,传递一种名为 "Eat-me signals" 的信号,吸引巨噬细胞来吞噬细胞。
- 参与凝血级联反应,PS的外翻使得血小板膜外表面负电荷聚集,成为凝血因子复合物的高效组装平台。
更多内容可以参考以下链接!
Eat-me signals - wikipedia: https://en.wikipedia.org/wiki/Eat-me_signals
Flippase - wikipedia: https://en.wikipedia.org/wiki/Flippase
Scramblase - wikipedia: https://en.wikipedia.org/wiki/Phospholipid_scramblase
The ins and outs of phospholipid asymmetry in the plasma membrane: roles in health and disease
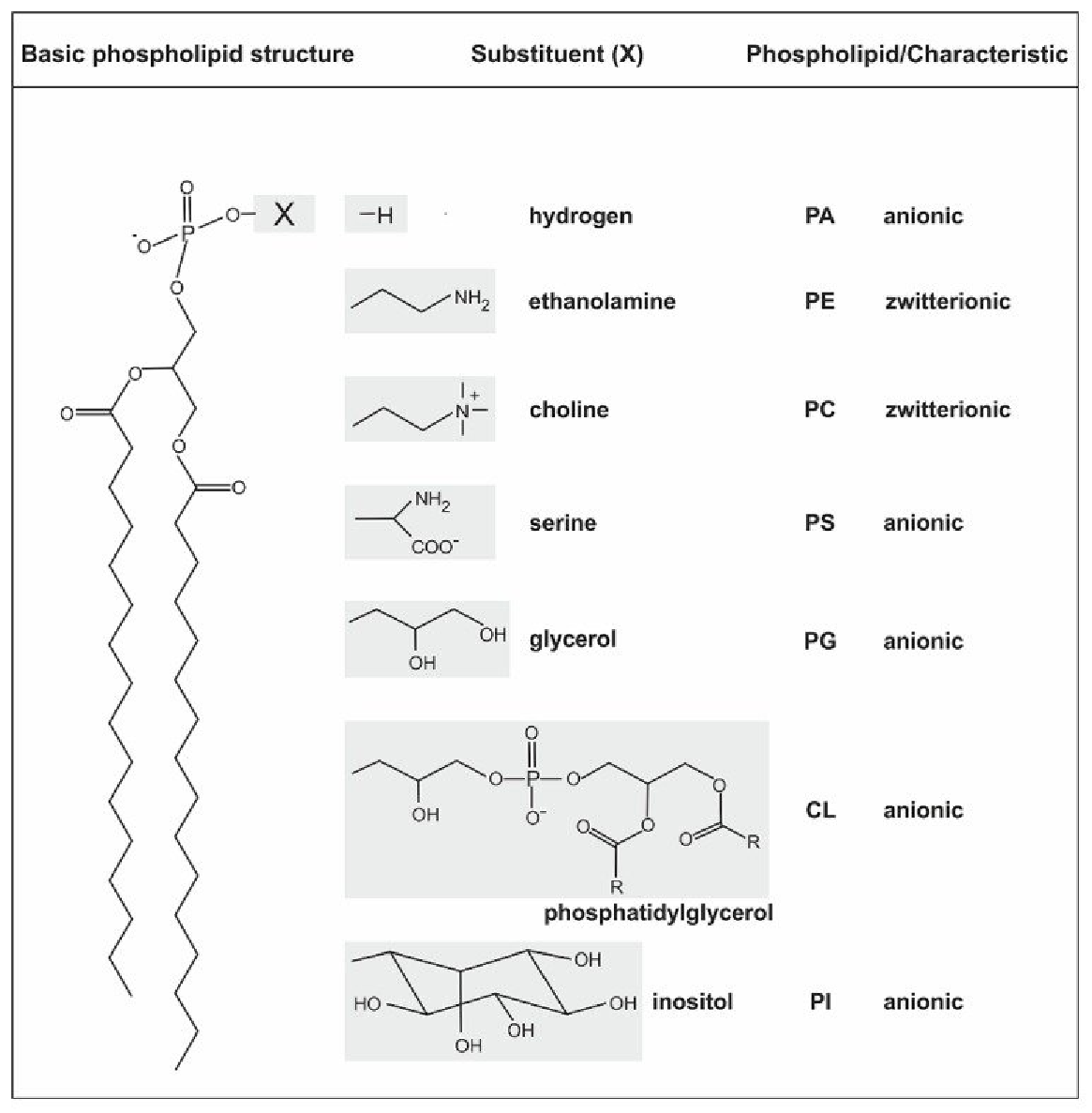
生物膜上常见磷脂种类的总结如下:
- 磷酸甘油酯/甘油磷脂
- 卵磷脂(磷脂酰胆碱,PC)
- 脑磷脂(磷脂酰乙醇胺,PE)
- 磷脂酰丝氨酸(PS) - 磷脂酰肌醇(PI) - 心磷脂(二磷脂酰甘油,CL) - 鞘磷脂(SM)
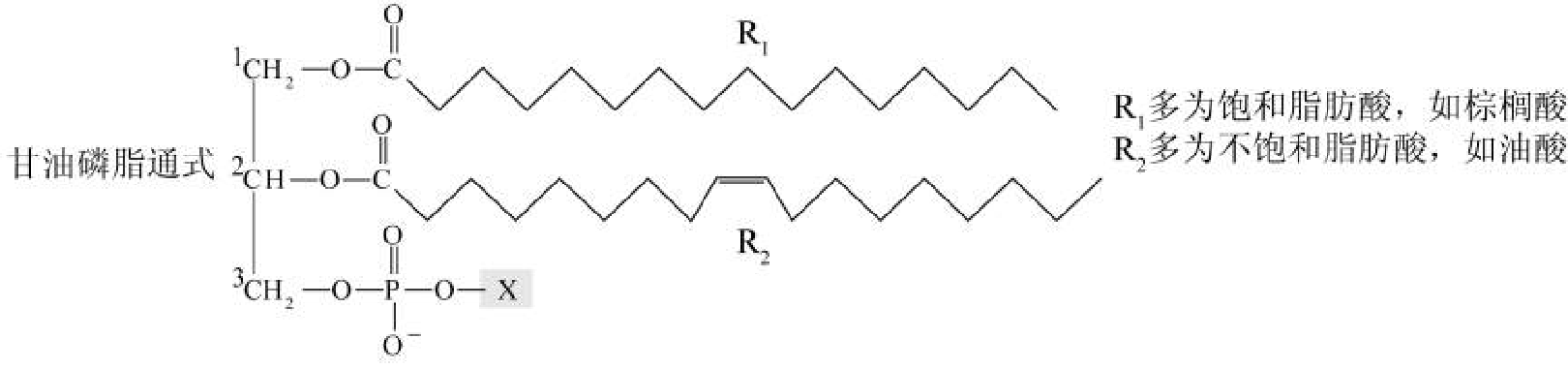
甘油磷脂
组成:甘油、脂酸、磷脂、含氮化合物。
结构: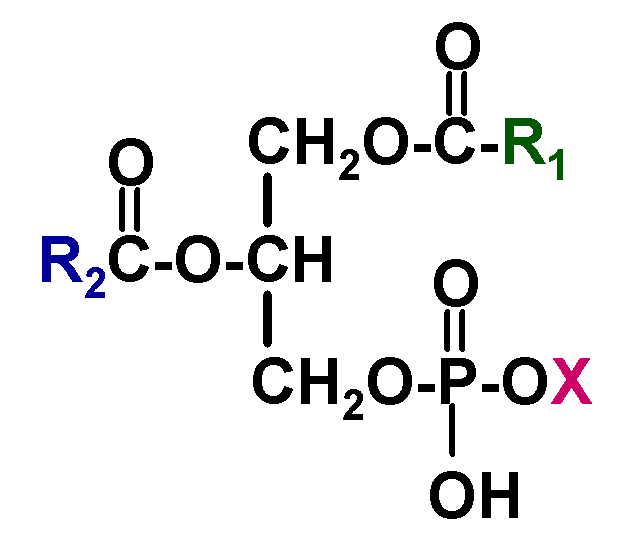
- \(\ce{R}\)为不饱和脂肪酸,常为花生四烯酸。
- \(\ce{X}\) = 胆碱、水、乙醇胺、 丝氨酸、甘油、肌醇、磷脂酰甘油等。
甘油磷脂含一个极性头、两条疏水尾,构成生物膜的磷脂双分子层。
几类重要的甘油磷脂:

鞘磷脂
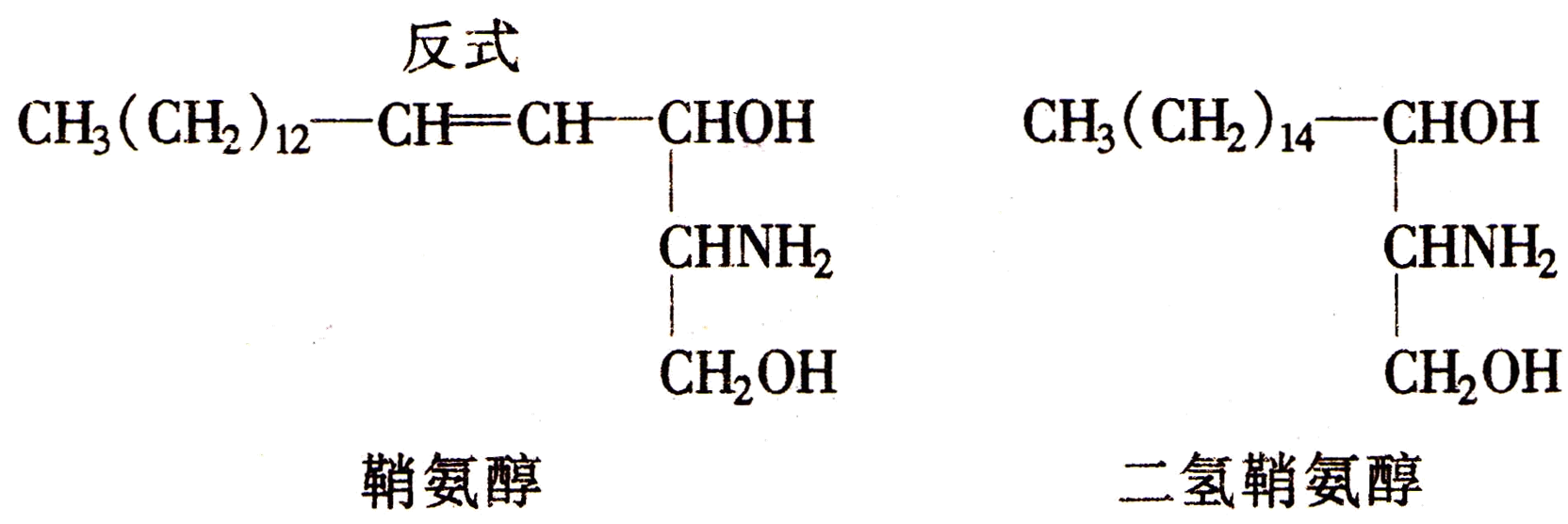
由鞘氨醇或二氢鞘氨醇构成的磷脂称为鞘磷酯。鞘氨醇和二氢鞘氨醇两者的结构如下:
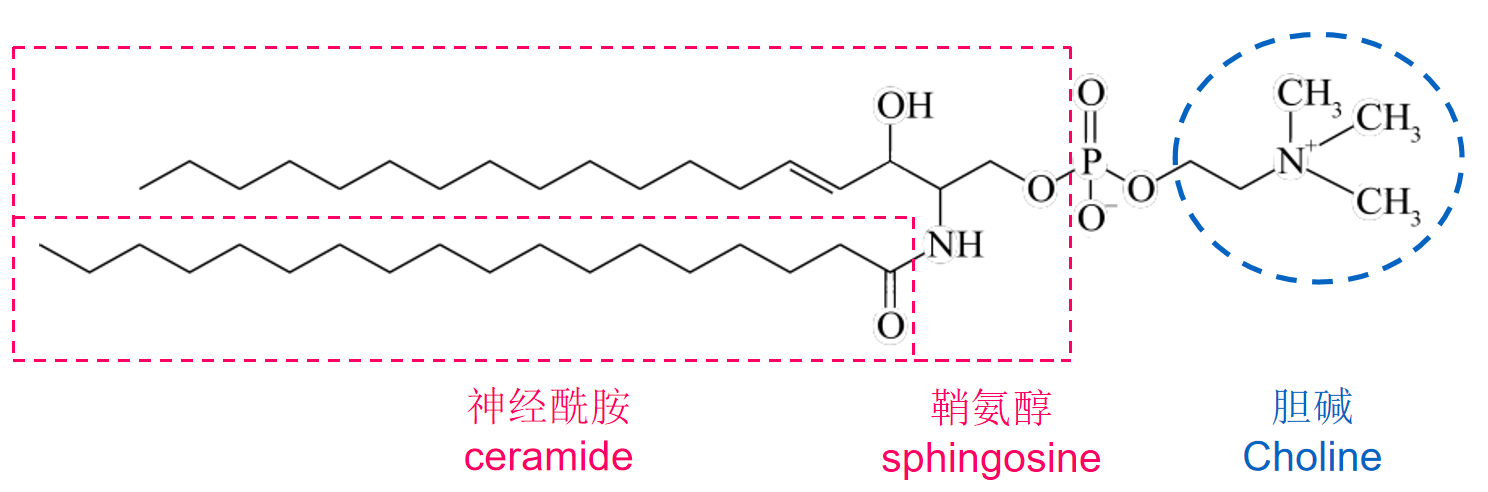
鞘氨醇磷脂(sphingophospholipids)的结构示意如下:
鞘氨醇-1-磷酸本身也是一种重要的信号脂类分子。
甘油磷脂和鞘磷脂的脂肪链长度不同,导致以鞘磷脂为主的双层膜较厚,或鞘磷脂的疏水长链深入膜脂双分子层的两个半叶之间,从而显著地增强膜的刚性和稳定性。
对于鞘磷脂,如果将其的磷酸基团替换为寡糖/单糖,则是鞘糖脂,鞘鞘磷脂和鞘糖脂都属于鞘脂。
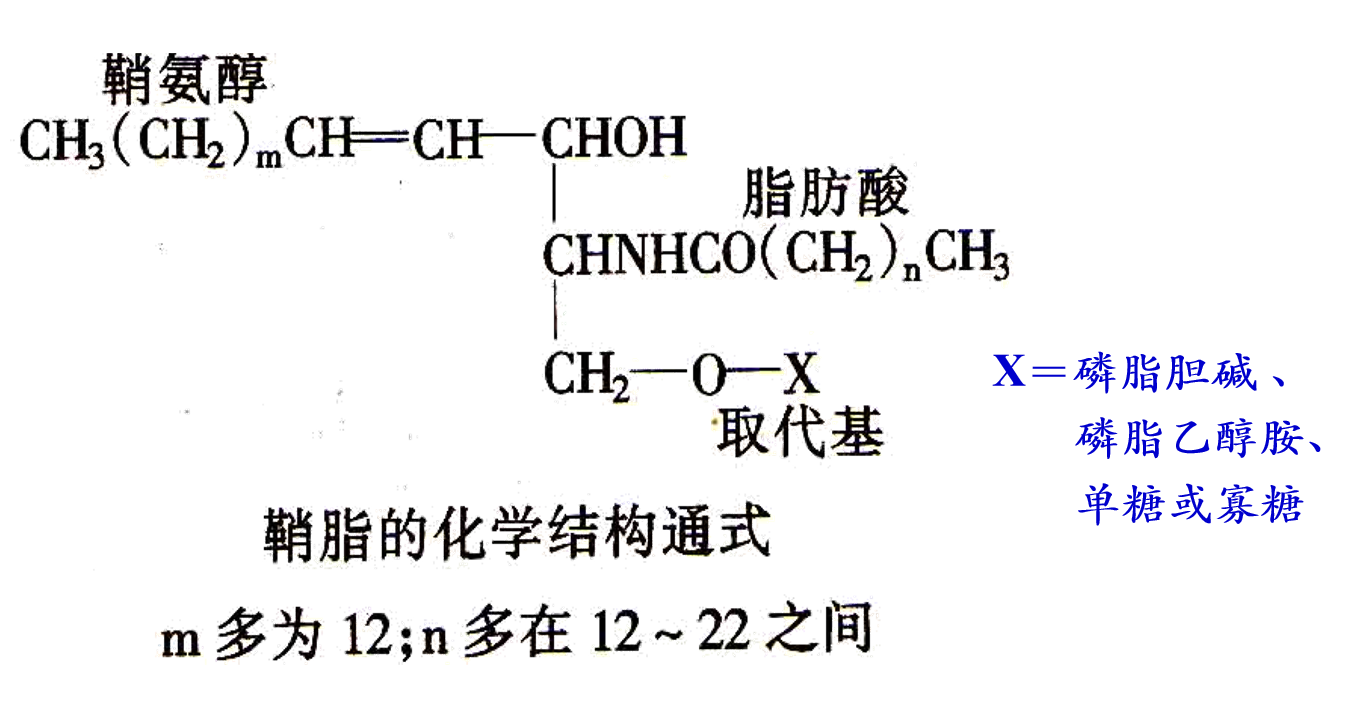
磷脂在体内具有重要的生理功能,包括有:
- 磷脂是构成生物膜的重要成分 - 卵磷脂存在于细胞膜中 - 心磷脂是线粒体膜的主要脂质
- 磷脂酰肌醇是第二信使的前体
- 缩醛磷脂存在于脑和心肌组织中
- 神经鞘磷脂和卵磷脂在神经髓鞘中含量较高
固醇
动物固醇
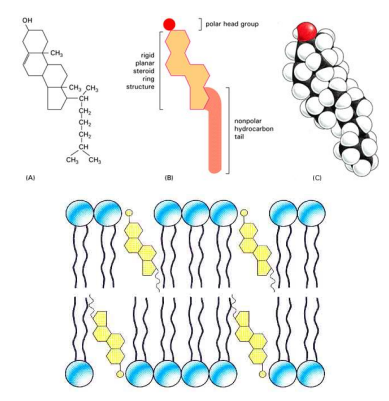
固醇在动植物中都有存在,动物固醇中最著名的是胆固醇,其是双亲性分子,在哺乳类的质膜中含量丰富。
胆固醇在体内分布广泛,主要存在形式有游离胆固醇、胆固醇酯,在肾上腺、卵巢等合成类固醇激素的腺体与脑等神经系统中的含量较高。
胆固醇对细胞膜的流动性、相行为、膜的分选与运输有调控作用,也可以直接对膜蛋白进行调控。
胆固醇上有类固醇环(甾环),其是刚性平面,可提高膜的刚性和微粘度;而疏水的分支脂肪烃链固有的运动性又增加膜局部微区的无序性,使膜流动性增加。
Note
类固醇对膜的这种双向调整和稳定作用使生物膜在较宽的温度范围(30-40℃)内行使功能。
动物体内可以自行合成胆固醇,其合成原料为乙酰CoA和NADPH,除成年动物脑组织及成熟红细胞外,几乎全身各组织均可合成,以肝、小肠为主。一般在细胞内的胞液、光面内质网上合成。
影响胆固醇合成的因素包括:
- 饥饿与饱食:饥饿可以显著抑制肝合成胆固醇,而高糖、高饱和脂肪膳食后胆固醇合成增加。
- 胆固醇:胆固醇自身可负反馈抑制肝胆固醇的合成。其主要抑制HMG-CoA还原酶的合成。
- 激素:胰岛素及甲状腺素,胰高血糖素及皮质醇两组分别促进/抑制HMG-CoA还原酶的合成,从而增加/减少胆固醇的合成。特别地,甲状腺素还促进胆固醇在肝转变为胆汁酸。
胆固醇有三个下游代谢途径:
- 在肝细胞中转化为胆汁酸,随胆汁经胆管排入十二指肠(主要)。
- 转化为类固醇激素。
- 转化为维生素\(D_3\)前体。
植物固醇
常见的植物固醇为豆固醇(stigmasterol)和谷固醇(sitosterol)。
糖脂
糖脂与磷脂有几乎完全相同的结构形式,也是二酰甘油或N-酰基鞘氨醇衍生产物,所不同的是亲水部分是由一系列单糖残基构成。包括以下几类:
- 甘油糖脂:)有MGDG(单半乳糖二酰基甘油),DGDG(双半乳糖二酰基甘油),SQDG(6-磺基-D-异鼠李糖二酰基甘油),叶绿体被膜和类囊体膜中含量较高
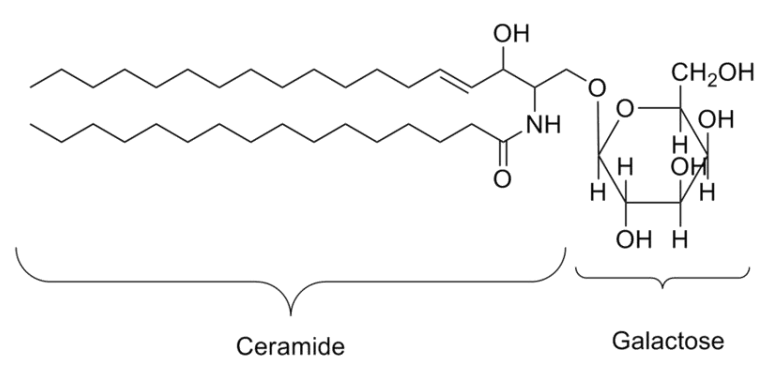
- 中性糖鞘脂:有半乳糖脑苷脂(galactocerebroside),脑苷脂是动植物细胞膜重主要的鞘脂,一般定为于脂双层的外叶。
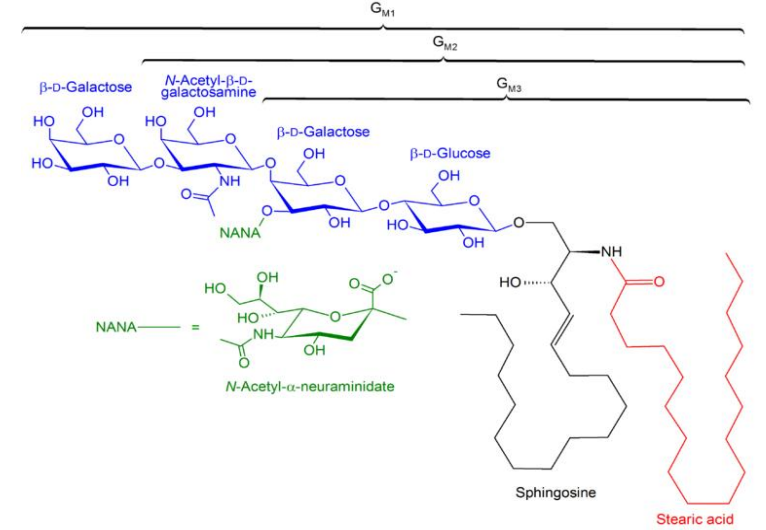
- 酸性糖鞘脂:有神经节苷脂(gangliosides, GM),其是最复杂的膜糖脂,约占神经细胞质膜总脂的5-10%。一些神经节苷脂还是某些毒素蛋白(破伤风杆菌和霍乱弧菌的外毒素)的受体。
膜蛋白
生物膜上的蛋白称为膜蛋白,其具有以下特点:
- 不同生物膜中蛋白质的种类和数量有很大差异。
- 生物膜的功能主要由膜蛋白承担。膜蛋白和膜脂分子共同维持膜的完整性、多样性和不对称性。
Note
膜蛋白的功能不仅取决于自身固有的结构,生物膜构成的特殊环境对膜蛋白形成并保持正确的构象起着不可或缺的作用。
膜蛋白与脂双层结合方式多种多样,按照不同的结合方式可分类为:
- 整合蛋白 (1, 2)
- 外周蛋白 (5, 6)
- 锚定蛋白 (3, 4)
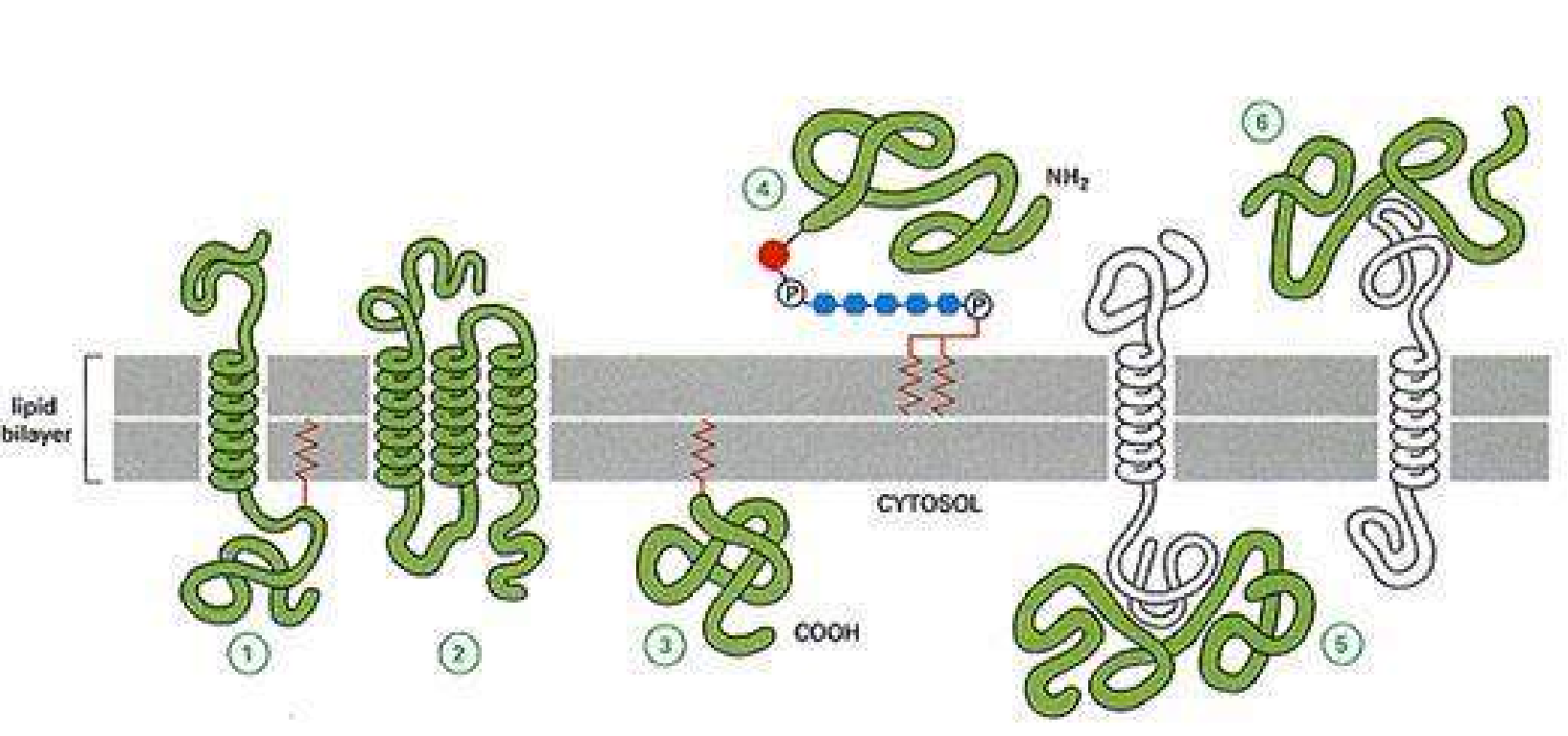
整合蛋白
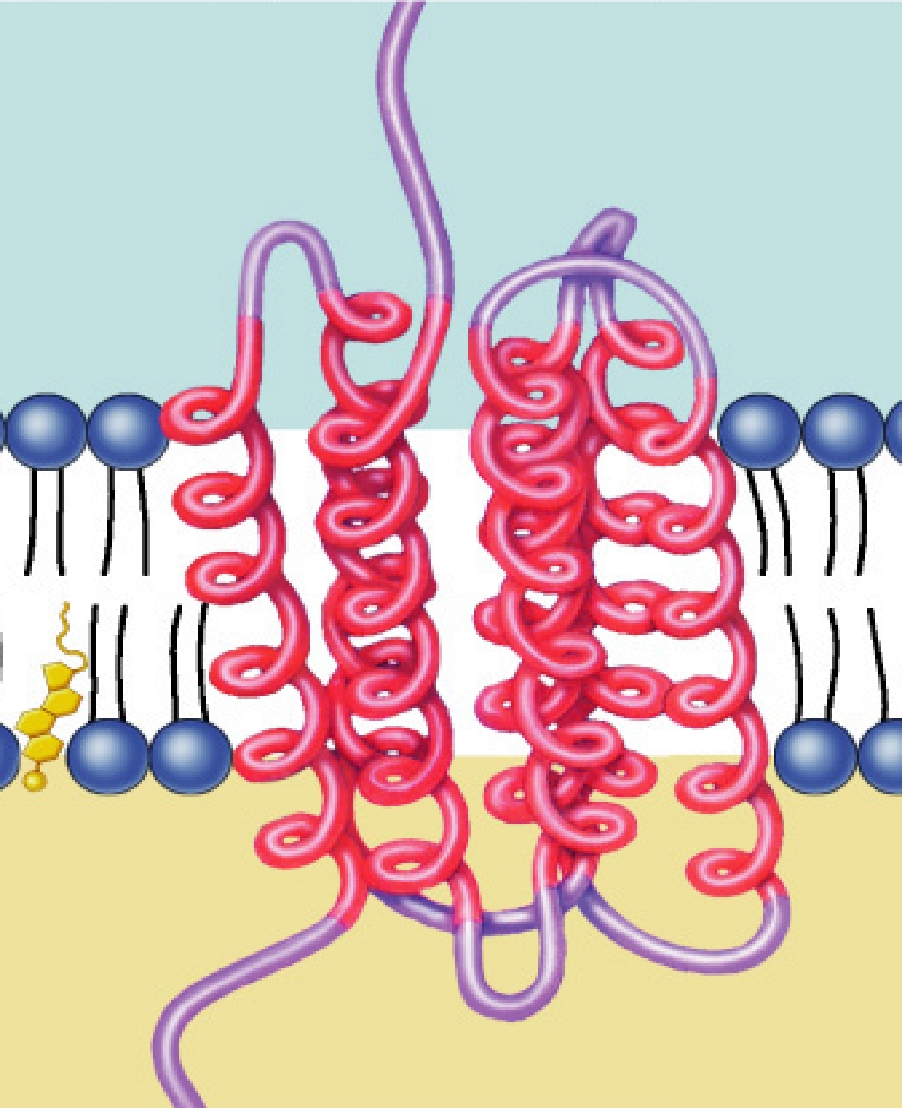
主要依靠疏水力与膜脂相结合,有的部分嵌在脂双层内(少数),有的横跨全膜,并且可能多次跨膜(大多数)。
整合蛋白较难分离,必须使用一些“雷霆手段”(去垢剂、有机溶剂和超声波)才能将其溶解下来。
整合蛋白具有胞外/胞内段与跨膜段,其中前者主要为亲水氨基酸,后者主要为疏水氨基酸,一个α螺旋组成的跨膜段大概占据20-30个氨基酸。
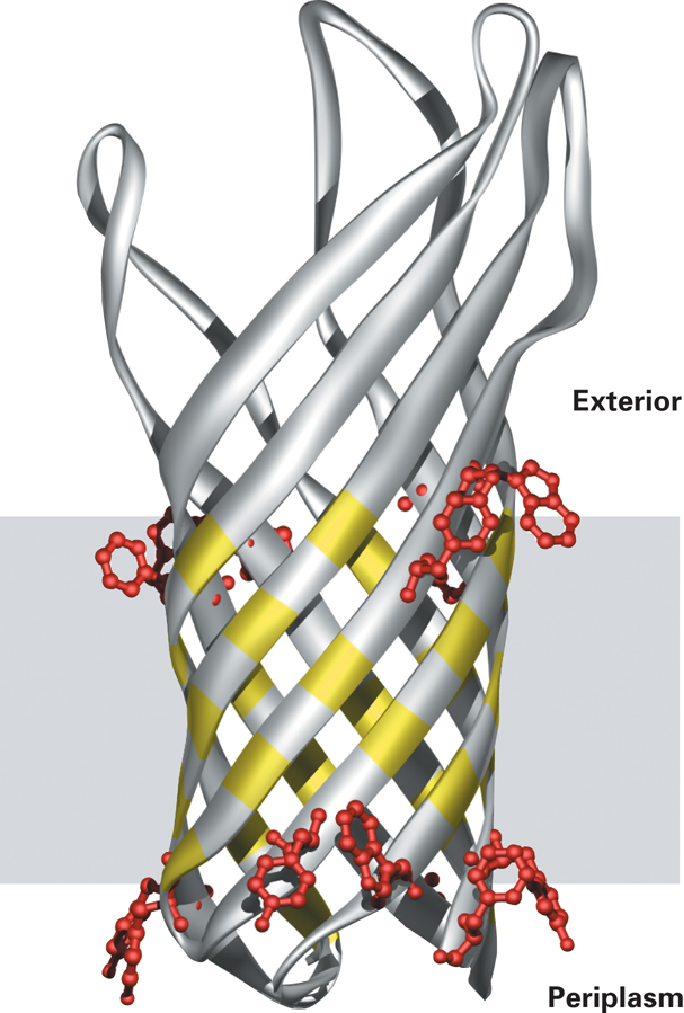
常见的跨膜段类型包括有α螺旋跨膜与β片层桶跨膜。
外周蛋白
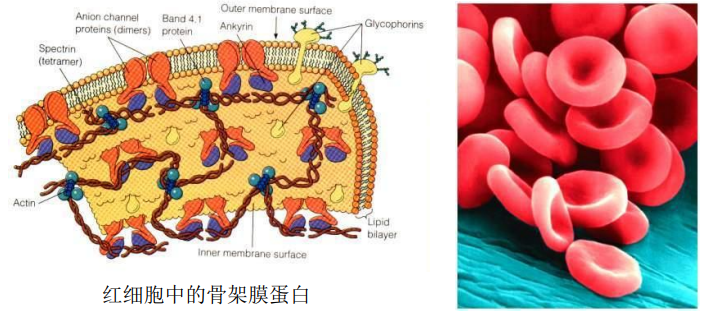
分布于膜的脂双层(外层或内层)的表面,通过静电力或非共价键与其他膜蛋白相互作用连接在膜上。
膜周边蛋白质比较易于分离,通过改变离子强度或加入金属螯合剂即可提取。
代表蛋白为红细胞中的骨架膜蛋白。
脂锚定蛋白
脂锚定蛋白(lipid-anchored) 又称脂连接蛋白(lipid-linked protein),通过共价健的方式同脂分子结合。同脂的结合有两种方式,一种是蛋白质直接结合于脂双分子层,另一种方式是蛋白并不直接同脂结合,而是通过一个或几个糖分子间接同脂结合。
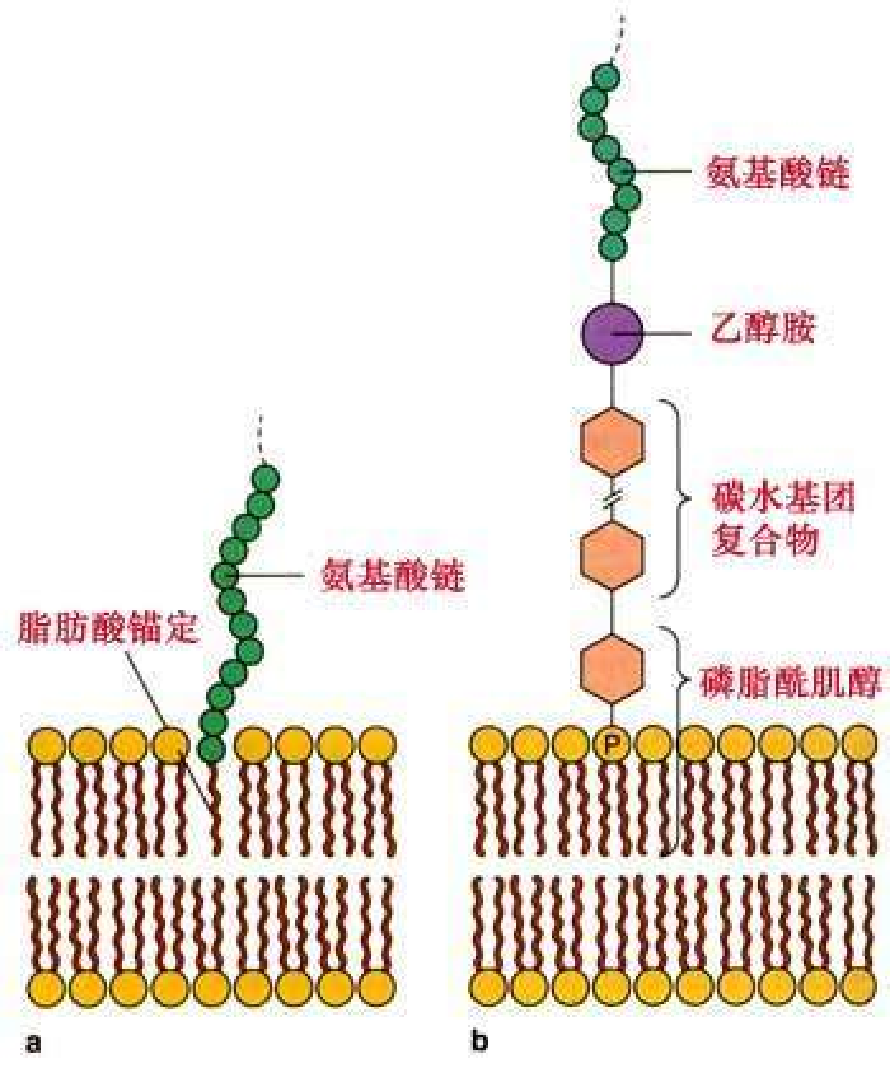
目前已知四种常见的脂锚定蛋白:
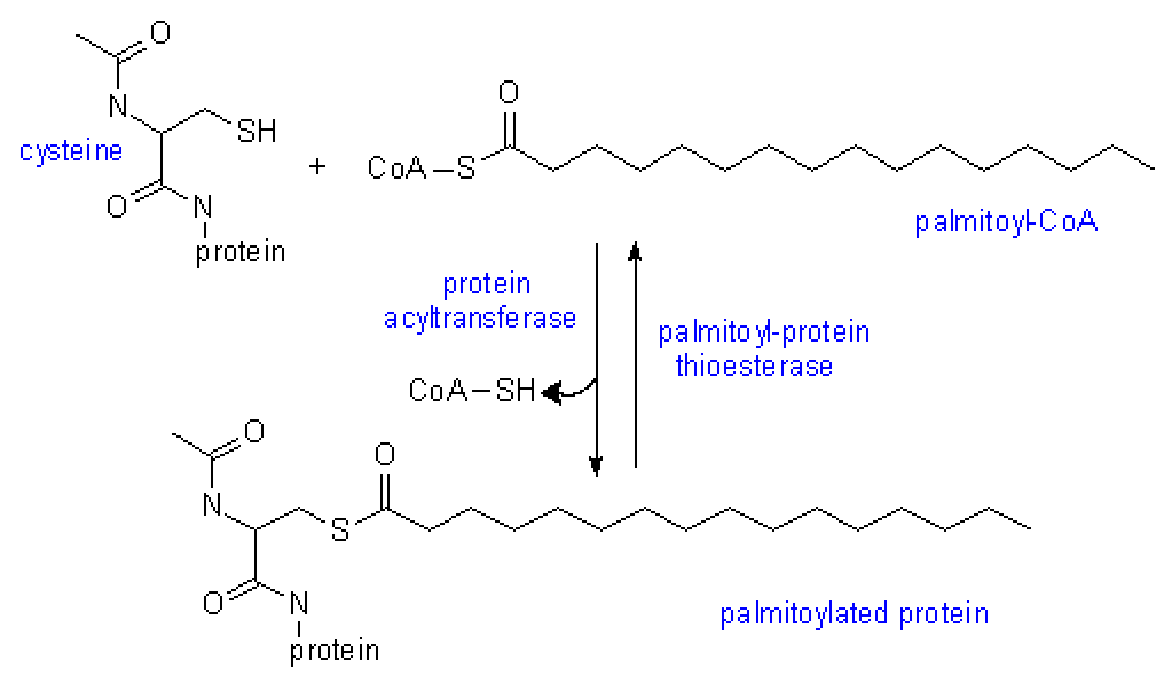
- 棕榈酰结合蛋白:肽链C-末端附近的Cys-SH残基与棕榈酰以硫酯键结合。
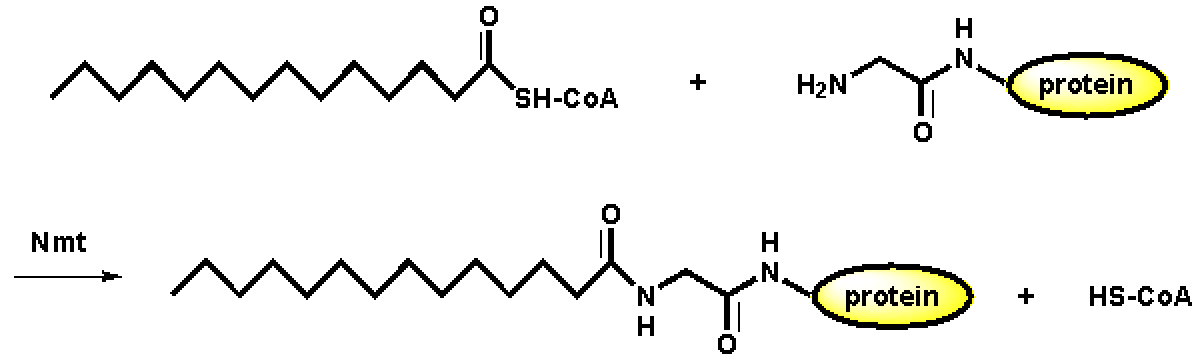
- 豆蔻酰结合蛋白:豆蔻酰结合于蛋白质的N-末端特定的氨基酸序列(1Gly-5Ser/Thr)。
- 异戊烯基化蛋白
- 糖基磷脂酰肌醇(GPI)锚定蛋白
膜糖
质膜含糖约占膜重2-10%,多为糖蛋白,少数为糖脂。构成生物膜的寡糖的单糖主要有9种:半乳糖、甘露糖、岩藻糖、葡萄糖、木糖、葡萄糖胺、N-乙酰葡萄糖胺、N-乙酰半乳糖胺和唾液酸。生物膜上的糖类几乎都定位于非胞浆面,在质膜外侧形成细胞外被或糖萼。这些成分可能是细胞表面受体,并且与细胞的抗原性有关。
膜蛋白和膜糖都具有不对称性:
- 膜蛋白的不对称性:膜蛋白的不对称性是指每种膜蛋白分子在细胞膜上都具有明确的方向性和分布的区域性。各种膜蛋白在膜上都有特定的分布区域。
- 糖的不对称性:无论在任何情况下,糖脂和糖蛋白只分布于细胞膜的外表面,这些成分可能是细胞表面受体,并且与细胞的抗原性有关。