细胞分裂与细胞周期
概述
我们首先要阐明细胞学的两大共识:
- 决定生物体大小的主要因素是细胞数量而不是细胞大小
- 新细胞是由旧细胞分裂产生的
细胞通过细胞分裂增殖,其中涉及两个概念:
- 细胞分裂:一个细胞分裂为两个子细胞的过程
- 分裂方式:有丝分裂与无丝分裂
- 细胞周期:从上次一分裂完成到下一次细胞分裂结束位置所经历的全过程。
Note
细胞分裂的生物学意义在于参与新个体的产生、器官的形成与替换死亡的细胞。
无丝分裂
无丝分裂,又称为二元分裂,最早发现于鸡胚红细胞。
无丝分裂过程中间期的细胞核拉长成哑铃状,中央部分变细断开,细胞随之分裂成两个。无丝分裂不形成纺锤丝,也不形成染色体,因此分裂后遗传物质不一定平均分配给两个子细胞。
有丝分裂
是真核细胞体细胞最基本的分裂方式,其特征是分裂时期染色质形成丝状或带状的染色体,并形成由纺锤丝组成的纺锤体。其结果是将染色质等量地分配到两个子细胞,使亲代和子代细胞具有相等的遗传物质,以保证遗传性状的稳定性。
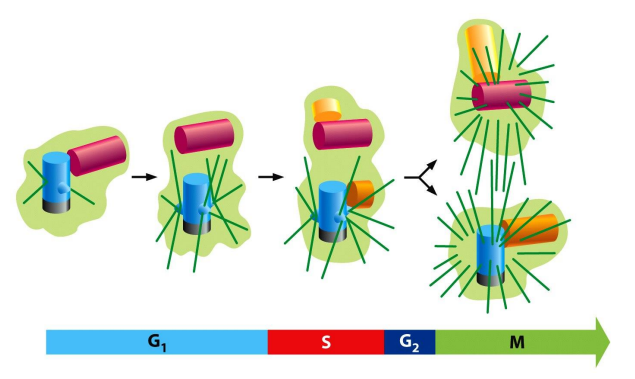
细胞周期概述
一个细胞周期从上一次分裂完成开始,到下一次分裂完成结束,包含四个阶段:
- G1期:DNA复制准备
- S期:DNA复制
- G2期:细胞分裂准备
- M期:细胞分裂,包含核分裂与细胞质分裂
最基本的两个事件:遗传物质的复制,遗传物质的分离与细胞的分裂。
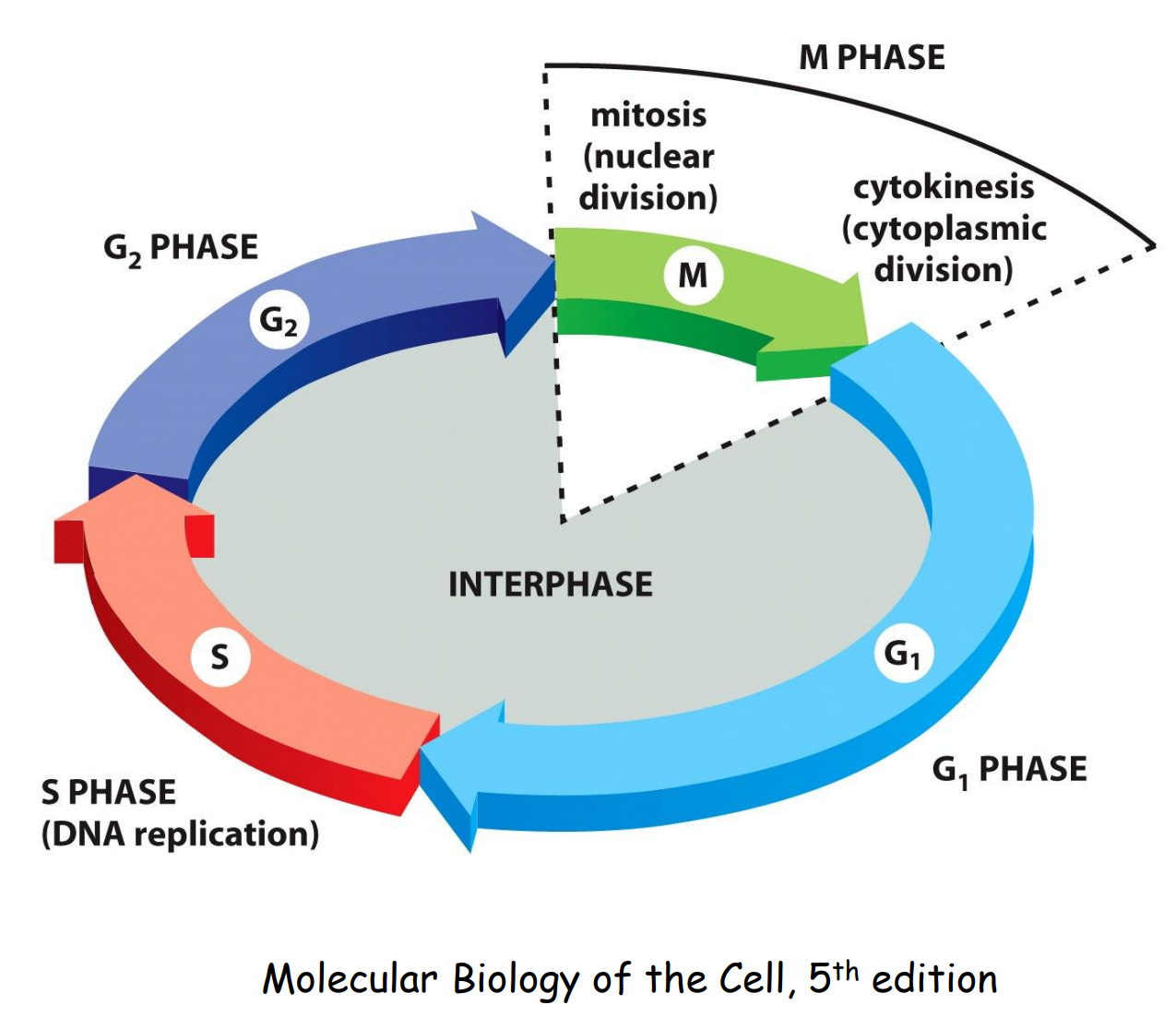
在所有阶段中,S期通常是耗时较长的,因为DNA复制是精密且困难的,但是应根据不同的细胞进行区分,比如受精卵在早期卵裂中通常不存在Gap期,直接以SM快速循环(受精卵有充足的营养物质,故早期无需准备与生长)。
Gap期的意义
Gap期相当于细胞在分裂中的一个准备,在Gap期中细胞可以准备营养物质,合成必要的蛋白。
Gap期与其他阶段的衔接往往也存在检查点,用于保证DNA复制和分裂的万无一失。
细胞生长主要发生在G1期、S期和G2期。
细胞周期
概述
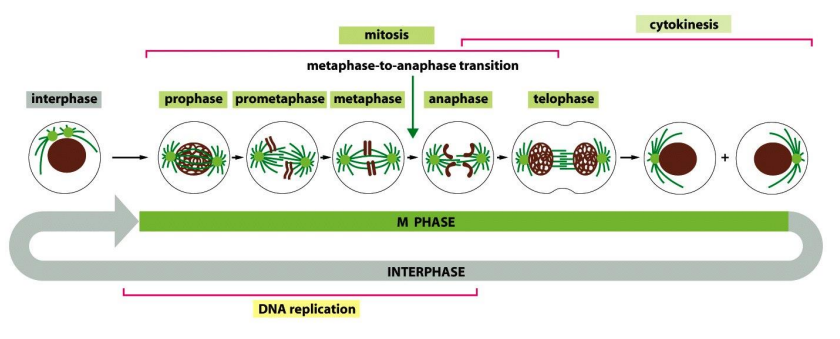
可见下图中,M期分为遗传物质的分配与胞质的分配两个阶段。
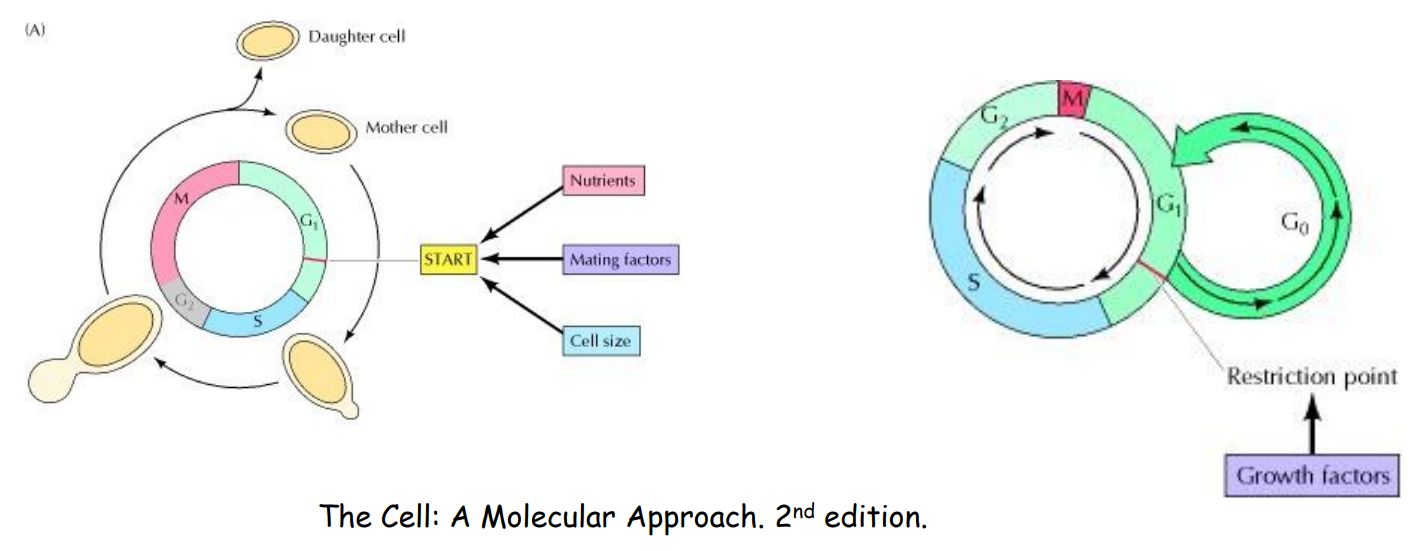
DNA合成期 (S phase)
动物细胞中,其决定是否进入S期由限制点 (Restriction Point) 把关。(在酵母中被称为 Start)
这个检查点通过了就不能反悔了!所以这个检查点非常关键。
细胞在S期的核心任务是完成DNA复制,有两个关键点:
- 确保遗传物质复制的高度保真性
- 确保基因组内所有区域被复制且仅复制一次
它的解决方法是:
- 使用起始识别复合物 (ORC) 结合特定DNA序列作为起始位点
- 在 ORC 基础上组装前复制复合物 (preRC)作为复制启动。
- 通过每周期仅形成一次的执照因子 (License) 保证复制只进行一次。
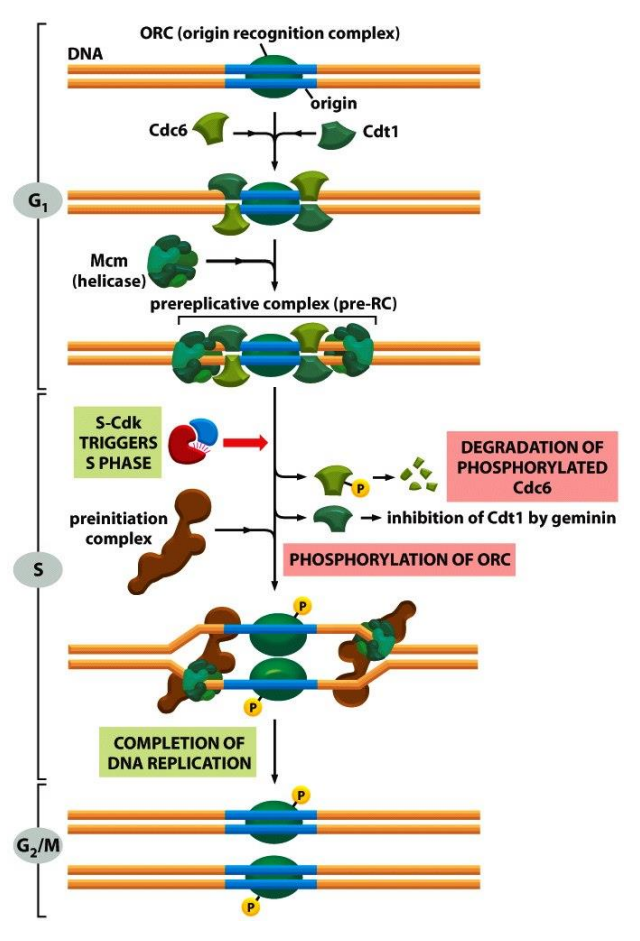
复制结束的DNA组装成紧靠在一起的姐妹染色体。
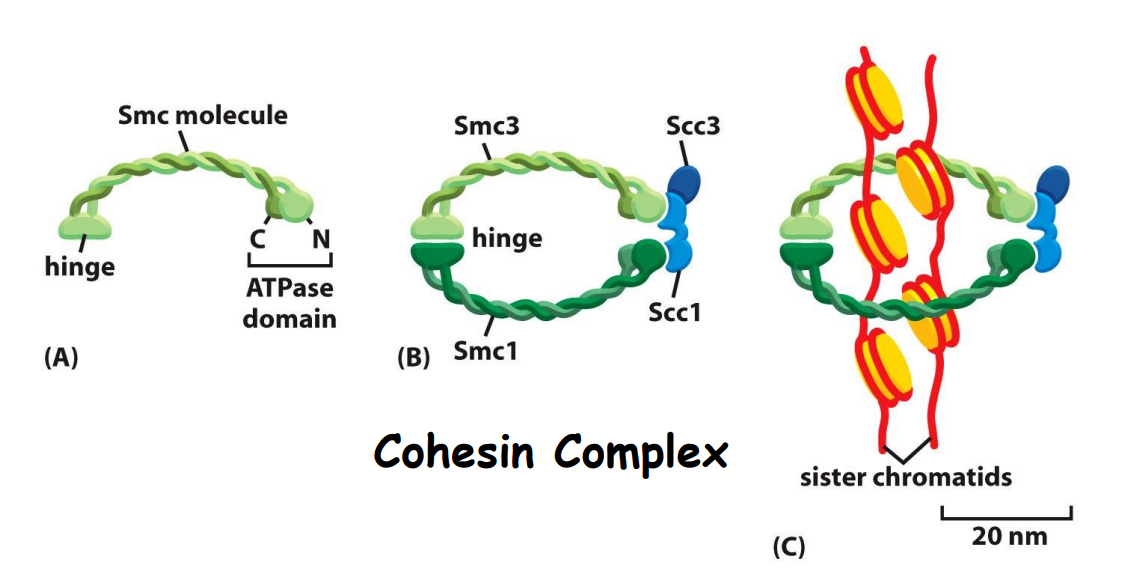
有丝分裂期
有丝分裂期可以进一步分为前期、前中期、中期、后期、末期、胞质分裂期。
有丝分裂前期
此阶段,染色质凝集、纺锤体开始形成,中心体在此过程中移向两极。
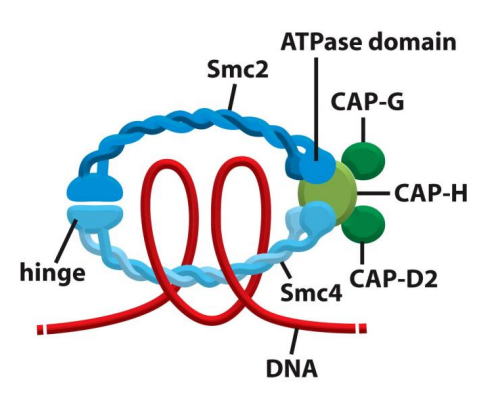
凝缩蛋白(Condensin)复合物协助染色质浓缩成为染色体。
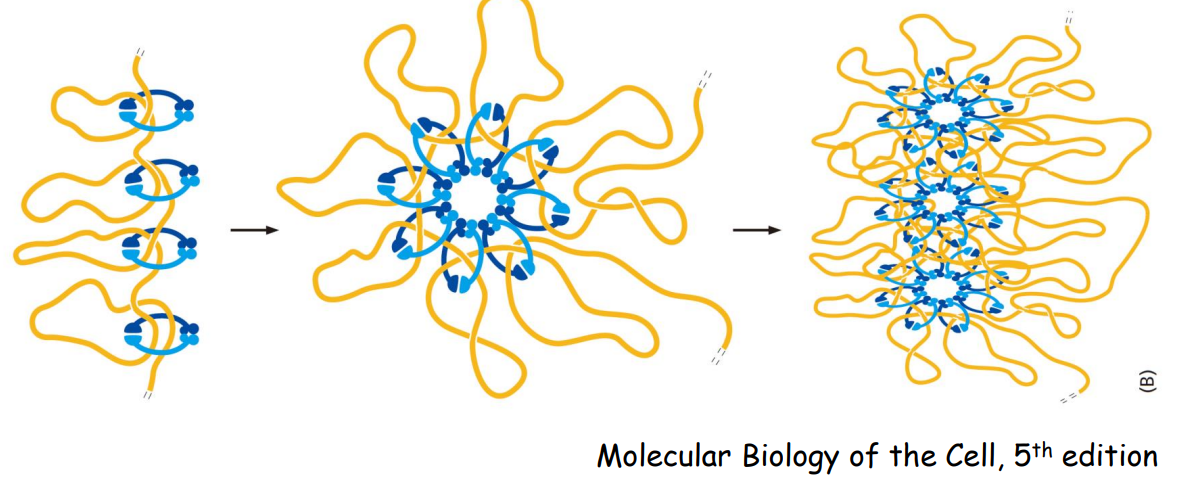
下图展示了纺锤体 (Mitotic Spindle) 的结构示意图:
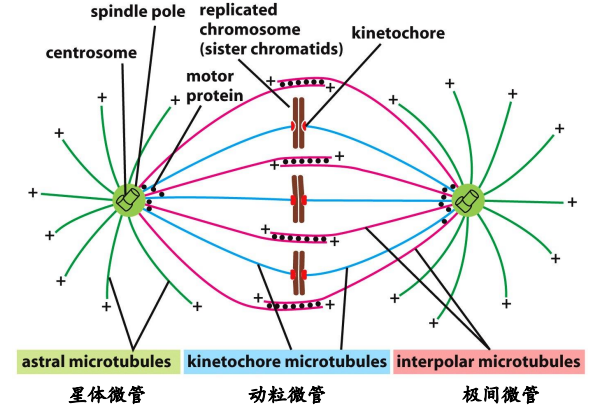
中心体
平日里细胞只有一个中心体,一个中心体有两个中心粒。
S期时候发生中心体的半保留形式。
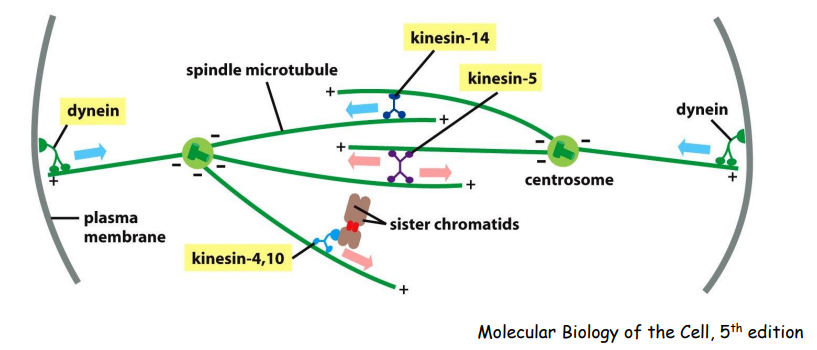
中心体的极向移动需要多种马达蛋白(驱动蛋白kinesins/动力蛋白dyneins)的参与。
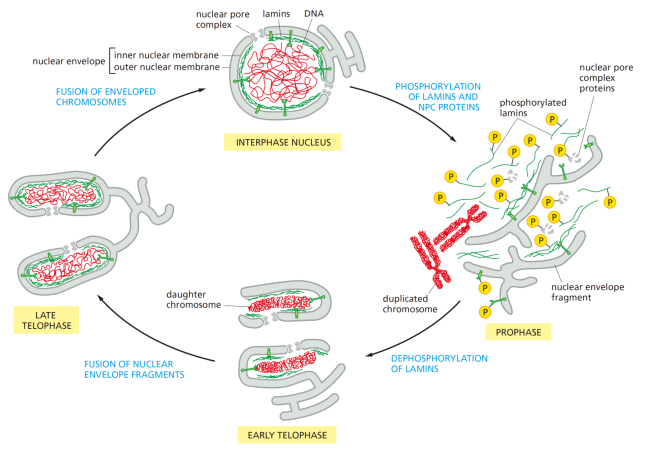
有丝分裂前中期
核膜崩解、纺锤体形成、染色体向赤道板运动,方便染色体被纺锤体捕捉。
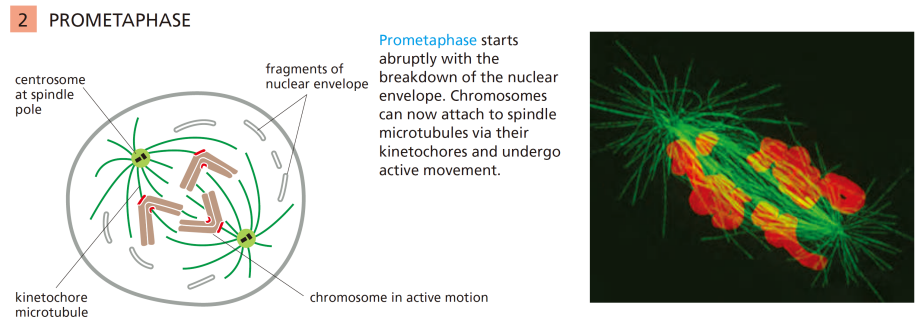
Note
核纤层磷酸化降解-核膜消失,与核纤层蛋白磷酸化有关。
有丝分裂极通过微管与染色体上的动粒相结合,从而捕捉染色体,使得染色体整齐排列在赤道板上。(非常耗能)
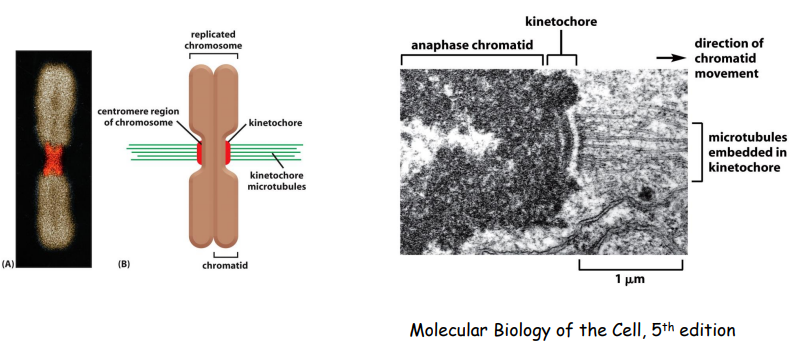
Note
游离于细胞质中的染色体不断振动,捕捉纺锤体微管,如果一条染色体两侧的姊妹着丝粒分别与两极来的微管相连,两侧的纺锤丝对染色体产生拉力,两侧拉力达到平衡时,染色体得以稳定排列在赤道面上。
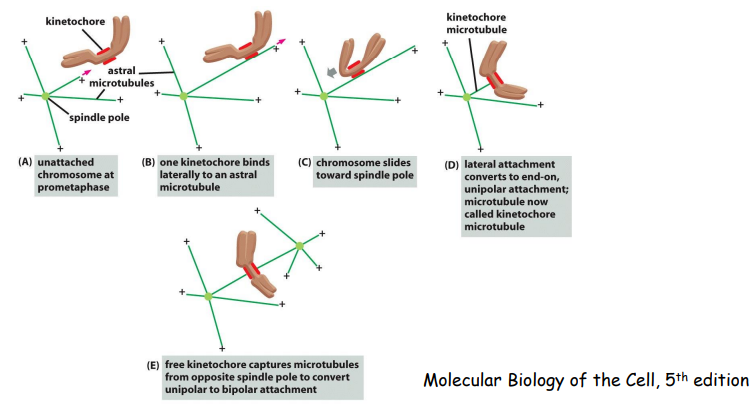
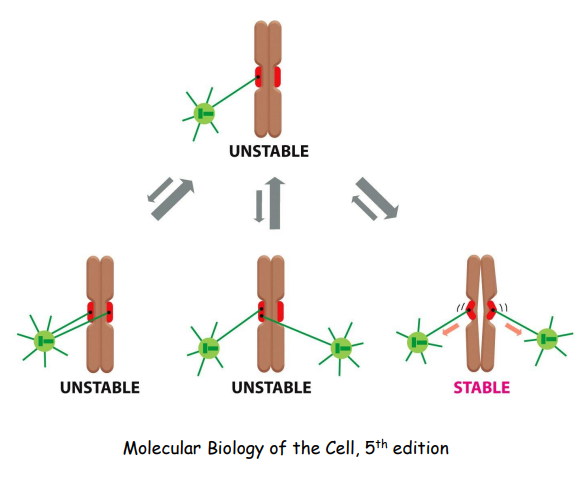
有丝分裂中期
染色体整齐排列到赤道板上:
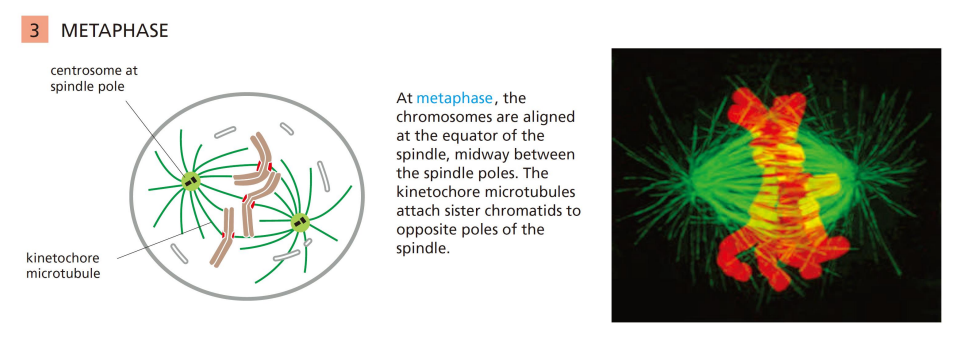
使得染色体整齐排列在赤道板 (metaphase plate) 上的张力来源于:
- 由动粒产生的向极拉力 (Ndc80复合物)
- 由动粒微管的微管流 (microtubule flux) 产生的向极拉力
- 极间微管之间的排斥推力(polar wind)
有丝分裂后期
姐妹染色单体分离,子代染色体到达两极。
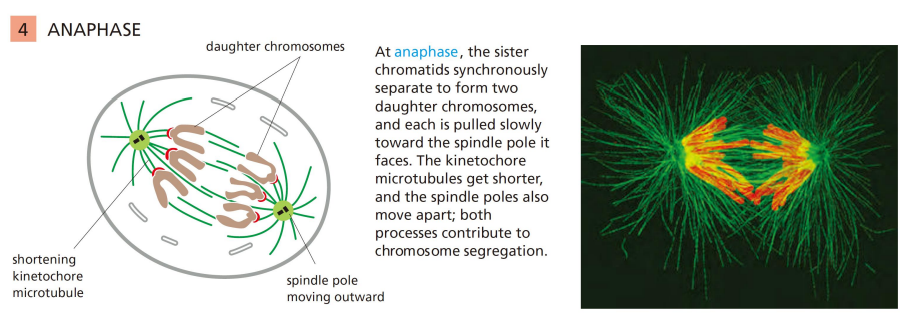
姐妹染色体的分离,首先是彼此间的连接 (cohesin) 的突然消失;其次由动粒与动粒微管连接处的向极拉力和马达蛋白移动所提供的牵引力共同完成分离。
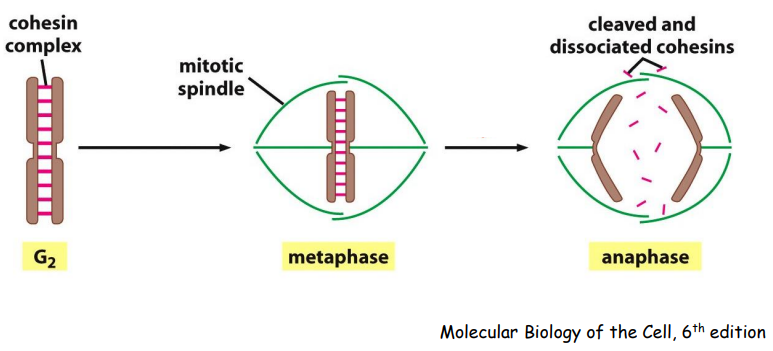
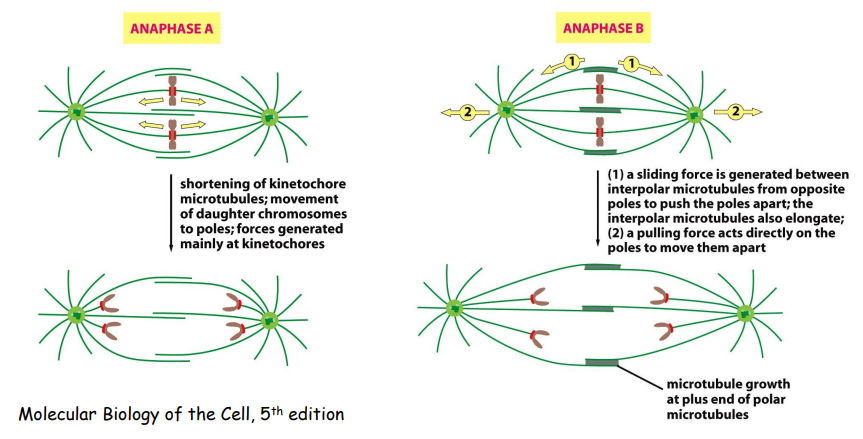
有丝分裂末期
该阶段形成子代细胞核:
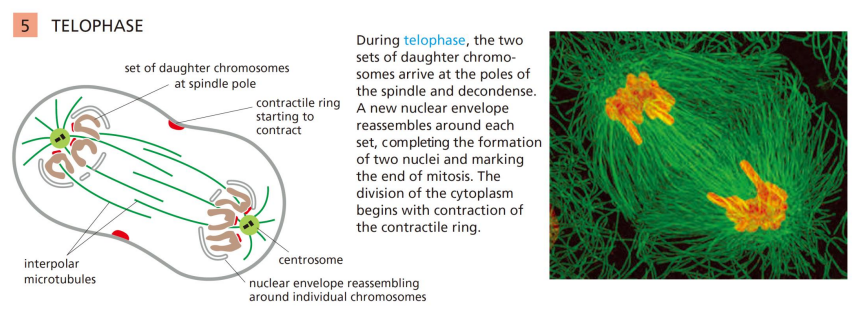
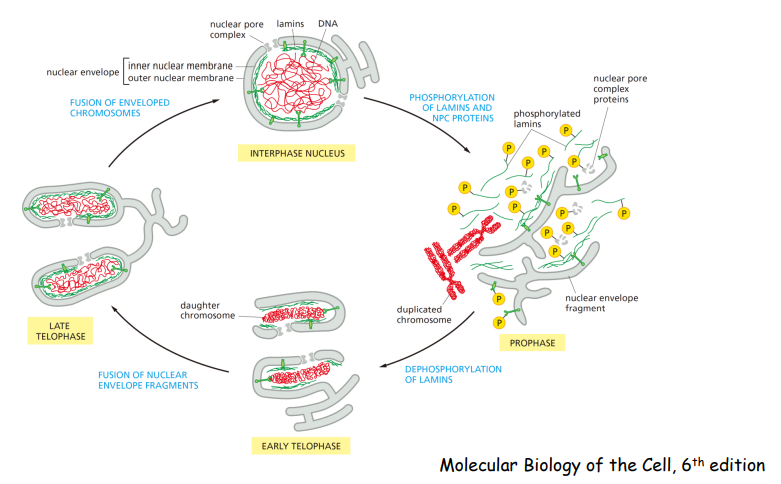
这过程核膜重建,重装子代细胞核:
胞质分裂期
胞质分裂期时收缩环和中体形成,细胞一分为二。
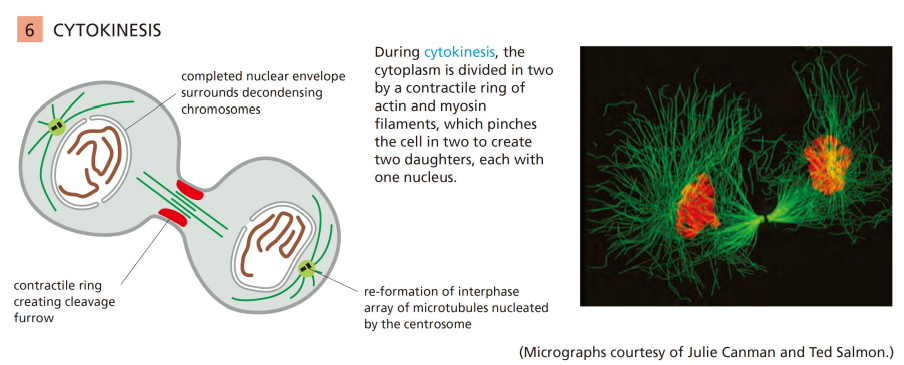
其中Actin和Myosin II介导收缩环的形成。
减数分裂
减数分裂发生于有性生殖的配子成熟过程中,其主要特征是DNA只复制一次,细胞连续分裂两次,所产生的子细胞中染色体数目比亲代细胞减少一半。
减数分裂对于维持生物世代间遗传的稳定性有重要意义。经减数分裂,有性生殖生物配子中的染色体数目减半,由2n变为n。经受精,配子融合形成的受精卵中染色体数又恢复为2n,由此保证了有性生殖的生物上下代在染色体数目上的恒定。
在减数分裂过程中发生染色体交换、重组等,促成了生物变异及多样性。
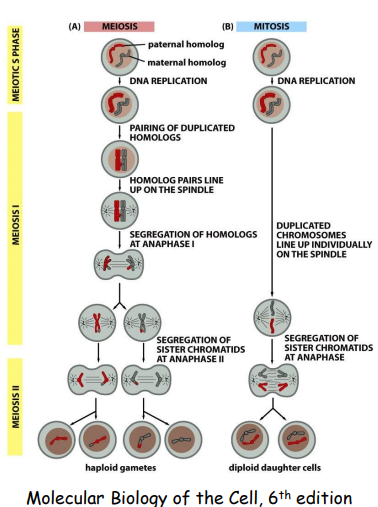
减数分裂中会发生一些重要的事件:
- Bivalent(二价体) 与 Chiasma(染色体交叉)
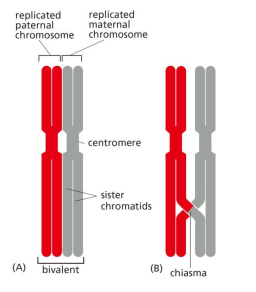
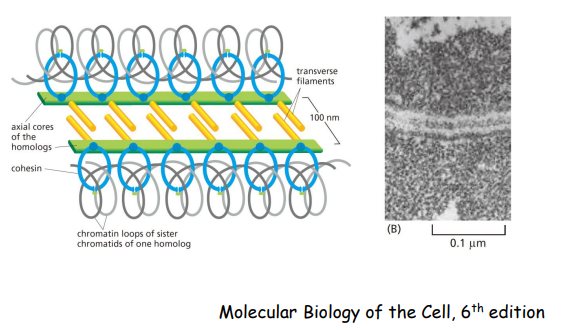
- Synapsis(联会)与Synaptonemal complex (联会复合体)
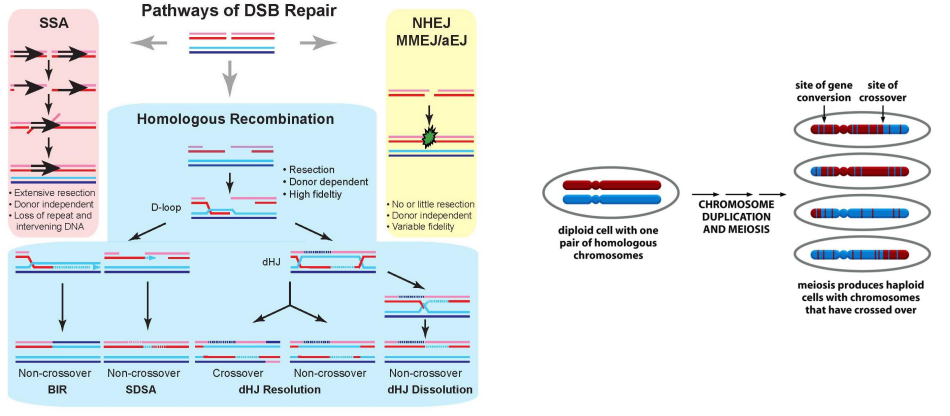
DSBR引起的crossover是减数分裂中同源染色体发生遗传物质交换的分子基础。
细胞周期调控
细胞周期驱动因素
细胞周期的关键调节因子是CDKs & cyclins:
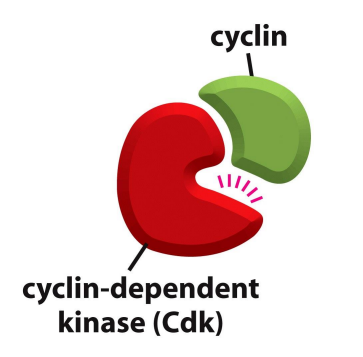
CDKs 是一种依赖于 cyclins 的蛋白激酶,CDKs 具有生物活性,被 cyclins 激活后可以磷酸化下游蛋白。
不同的 CDKs 在细胞周期中的量是少且恒定的,基本不会出现表达量的变化,其是否发生作用取决于其活性水平。
不同的 CDKs 与 cyclins 在不同的时期组合,在从而在适当时刻激发特定 CDKs 的活性,通过级联放大调控细胞周期的维持与推进。
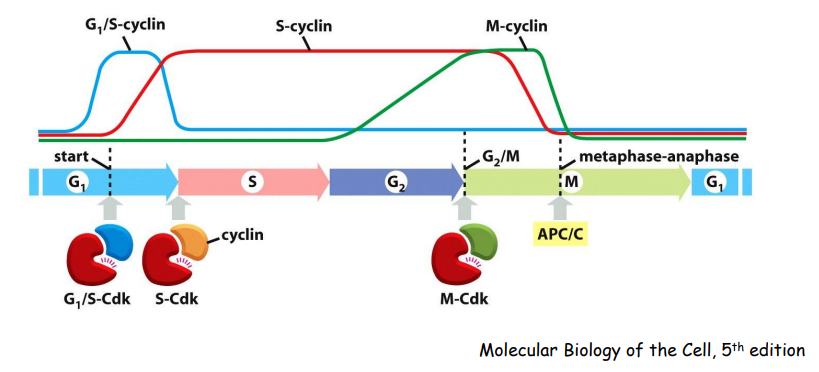
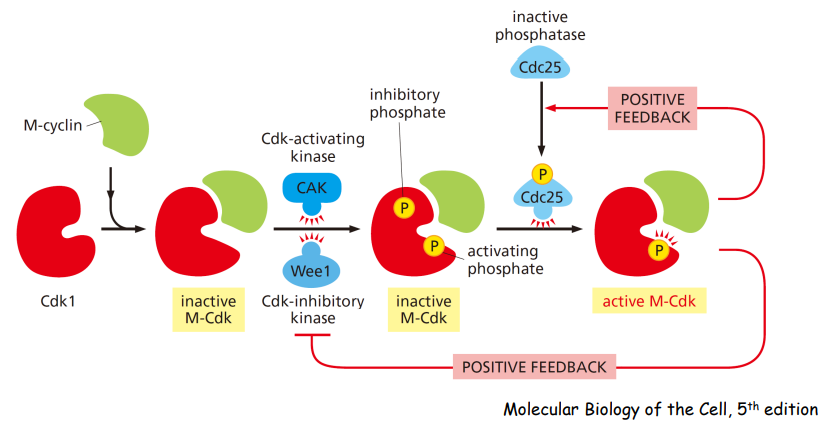
CDK活性调节的主要模式包括:
- Cyclin的结合
- Cdk-activating kinase (CAK)
- Inhibitory phosphorylation
- Cdk inhibitor proteins (CKIs)
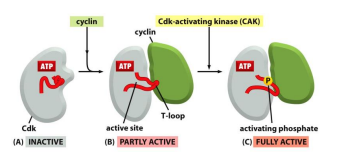
下图展示了 cyclins 的结合与 CDKs 自身被磷酸化从而激活活性的过程:
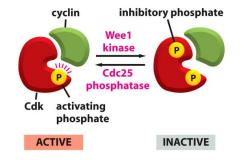
下图展示了 Wee1 kinase 通过磷酸化 CDKs 从而抑制其活性:
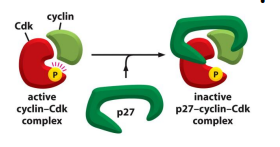
p27蛋白可以通过与 CDKs & cyclins 复合物结合来抑制 CDKs 的活性。
Mitogens (丝裂原,促进细胞分裂的物质,包括生长因子) 促进细胞进入有丝分裂:
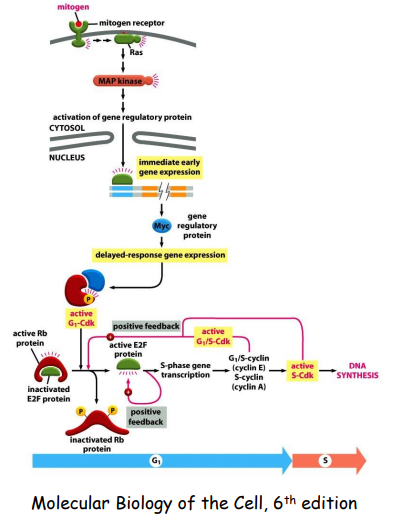
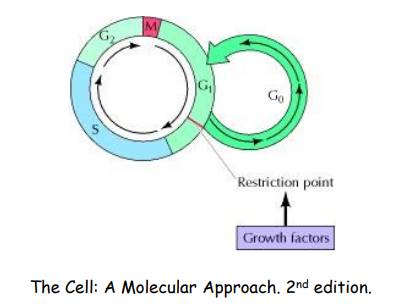
对于那些一直无法通过 G1 检查点的细胞,称为其为 G0 期的细胞:
在 M-CDK 的活化可以促进细胞进入 有丝分裂 (Mitosis):
G1->S的过程,一旦通过就无法刹车,而在之后的步骤中细胞可以随时停下来调整自己,而在有丝分裂过程中,有丝分裂中期和后期也有一个检查点,或者说 transition,它将检测细胞内的遗传物质是否正常,如果不正常可以随时暂停细胞的分裂甚至致其死亡。
而这个过程中 起到重要作用的就是后期促进复合物 APC/C,APC/C 的一个底物是 M-cyclins,因为其是促进细胞进入有丝分裂并维持在中期。APC/C 会促使其降解,推进有丝分裂向前。
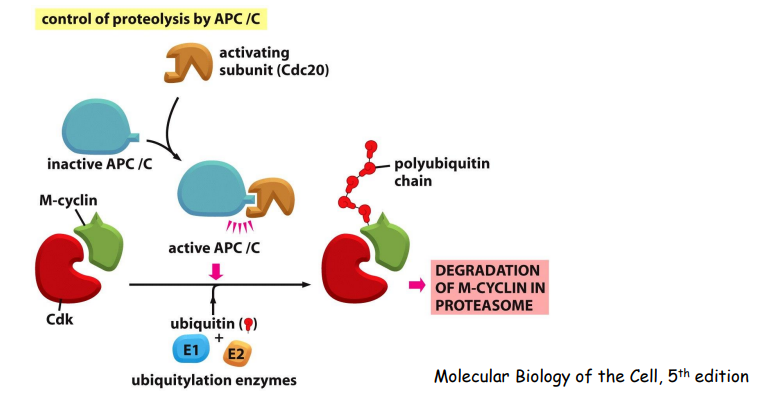
APC/C促进separase活化,而被活化的separase切割Cohesion complex,从而促进细胞向有丝分裂后期的转换.
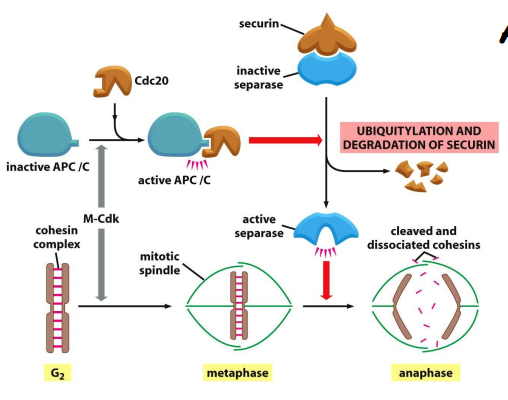
完成有丝分裂后,细胞还会维持一段时间 APC/C 复合物的活性,以使得 M-cyclins 蛋白量降低,避免细胞提早进入 M 期,得以在 G1 期休养生息一下。
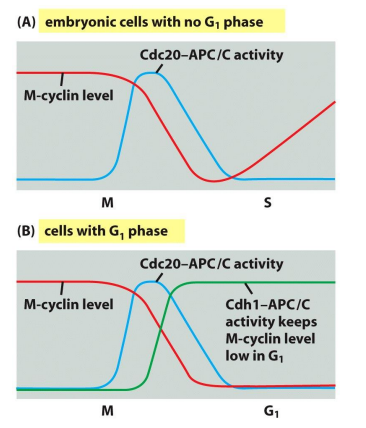
下图展示了整个细胞周期中,有丝分裂进程的关键调节分子:
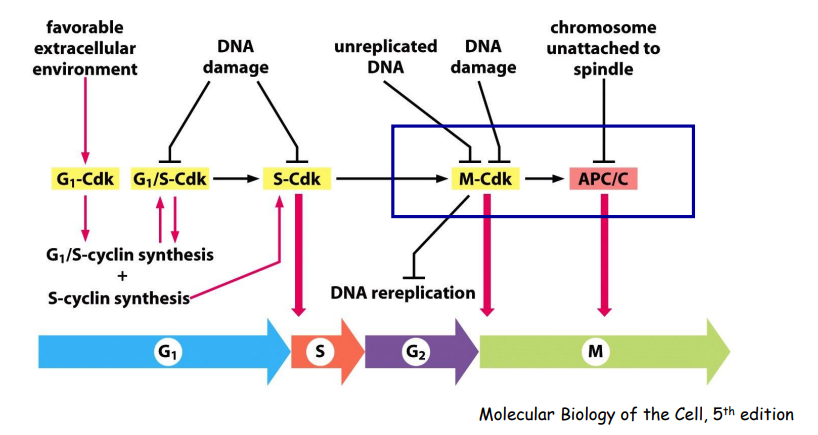
细胞周期监控方式
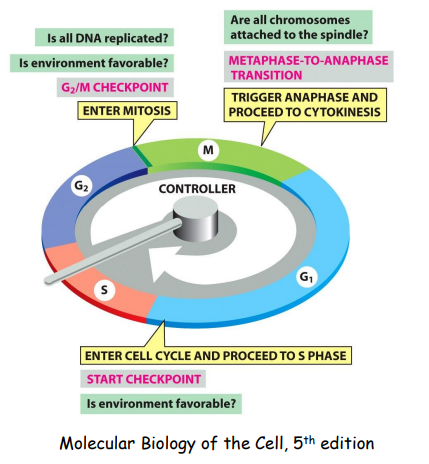
细胞周期检验点/检查点负责监控细胞周期进程,包含 G1/S、Intra S、G2/M和Spindle 四个检查点。
Intra S checkpoint
S期内检查点会实时监控DNA复制是否正常。
Note
一个典型的DNA损伤抑制细胞周期的模型如下:
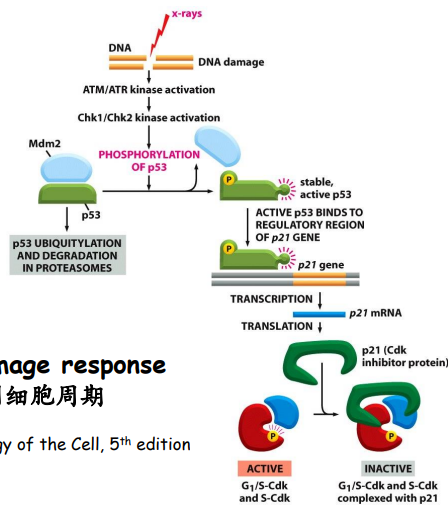
- Mitogens(有丝分裂原)促进细胞进入有丝分裂;
- Checkpoints(细胞周期检验点)监控周期进程;
- DNA damage response(DDR) 抑制细胞周期;
- 细胞的天然复制寿命决定了细胞能进入细胞周期的次数