核酸的结构和功能
核酸 (nucleic acid) 是以核苷酸 (nucleotide) 为基本组成单位的生物大分子,具有携带和传递遗传信息的功能。
核酸可以分为脱氧核糖核酸(deoxyribonucleic acid,DNA)和核糖核酸(ribonucleic acid,RNA)两类。
- 脱氧核糖核酸:简称为DNA,90%以上分布在细胞核,其余分布在线粒体内,携带遗传信息并通过复制传递给下一代。
- 核糖核酸:简称为RNA,分布在细胞核,细胞质和线粒体中,是DNA转录的产物,参与遗传物质的复制与表达。(某些病毒的RNA也可以承载遗传信息)
核酸的化学组成与一级结构
核苷酸的化学组成
DNA的基本组成单位是脱氧核糖核苷酸 (deoxyribonucleotide) ,RNA的基本组成单位是核糖核苷酸 (ribonucleotide) 。
碱基的主要作用是为了配对,可以认为它们是承载遗传信息的重要接口。
体内的嘧啶碱基有三种:尿嘧啶、胸腺嘧啶和胞嘧啶。
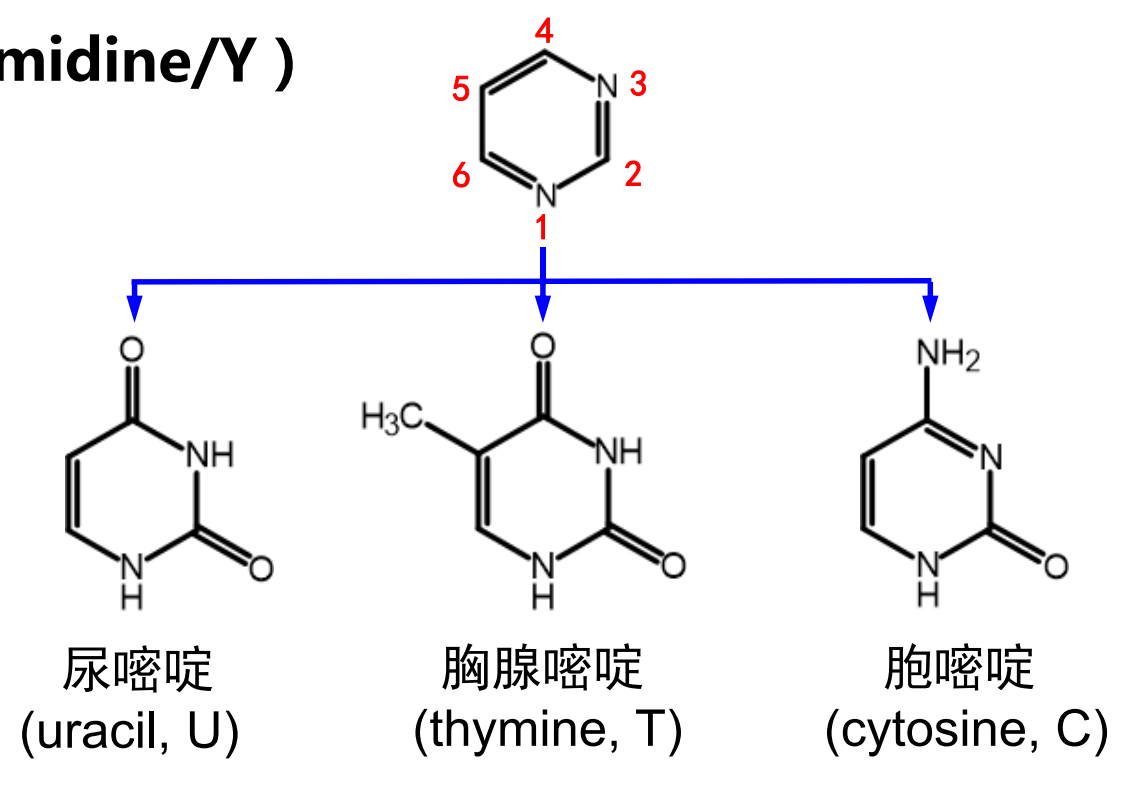
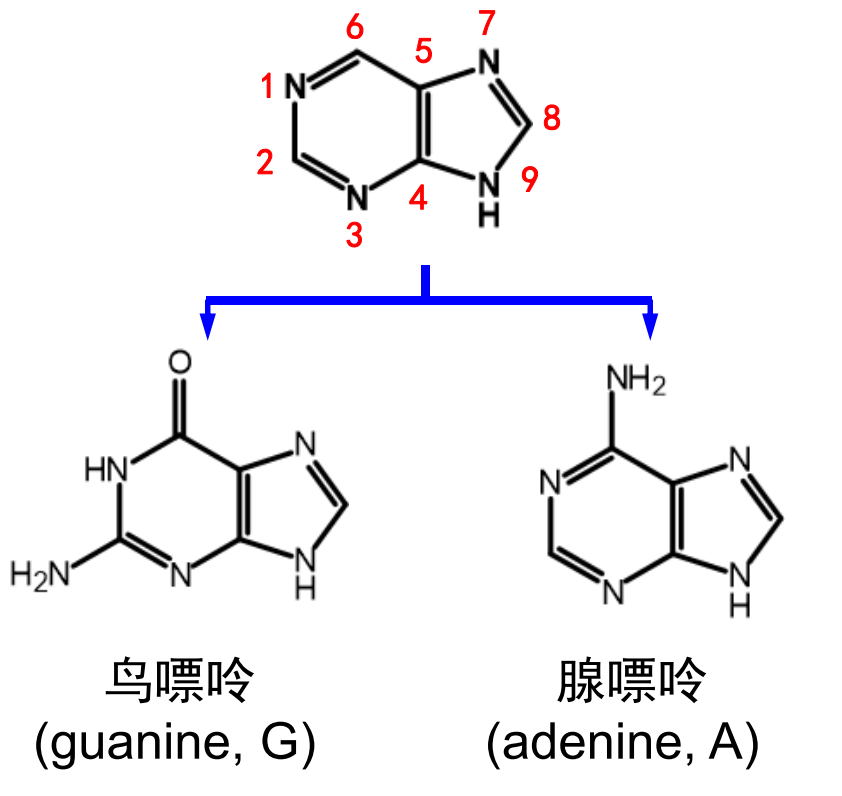
嘌呤碱基有两种:鸟嘌呤和腺嘌呤。
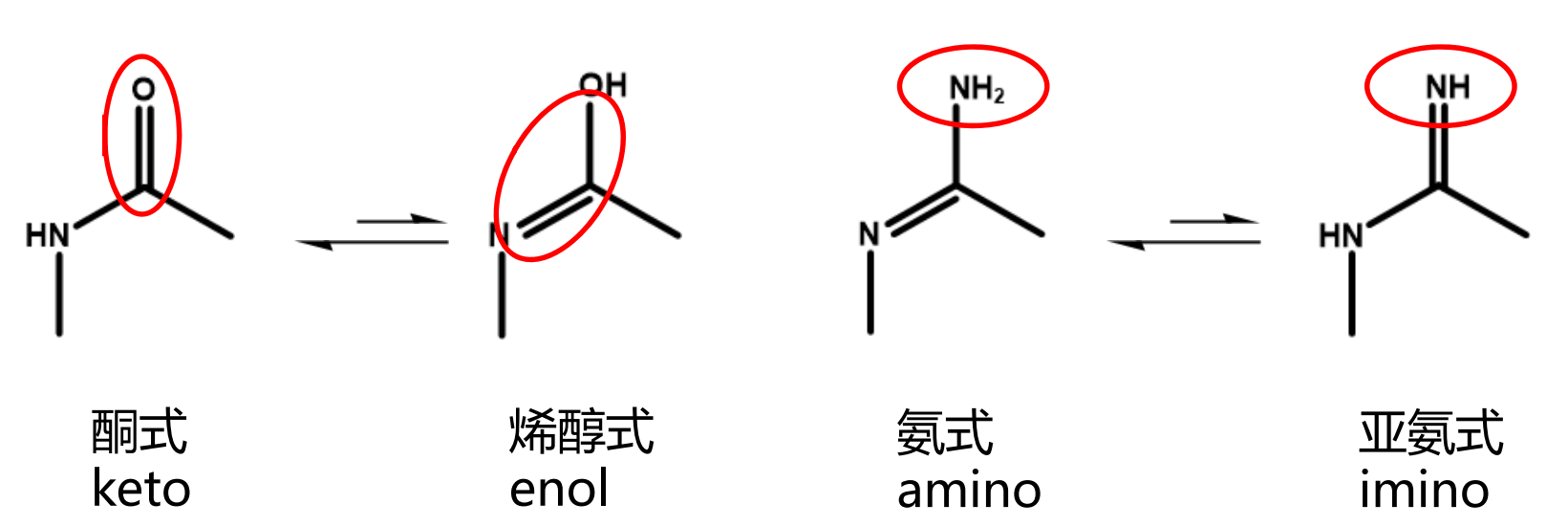
碱基存在互变异构体,其为氢键的形成奠定了基础。
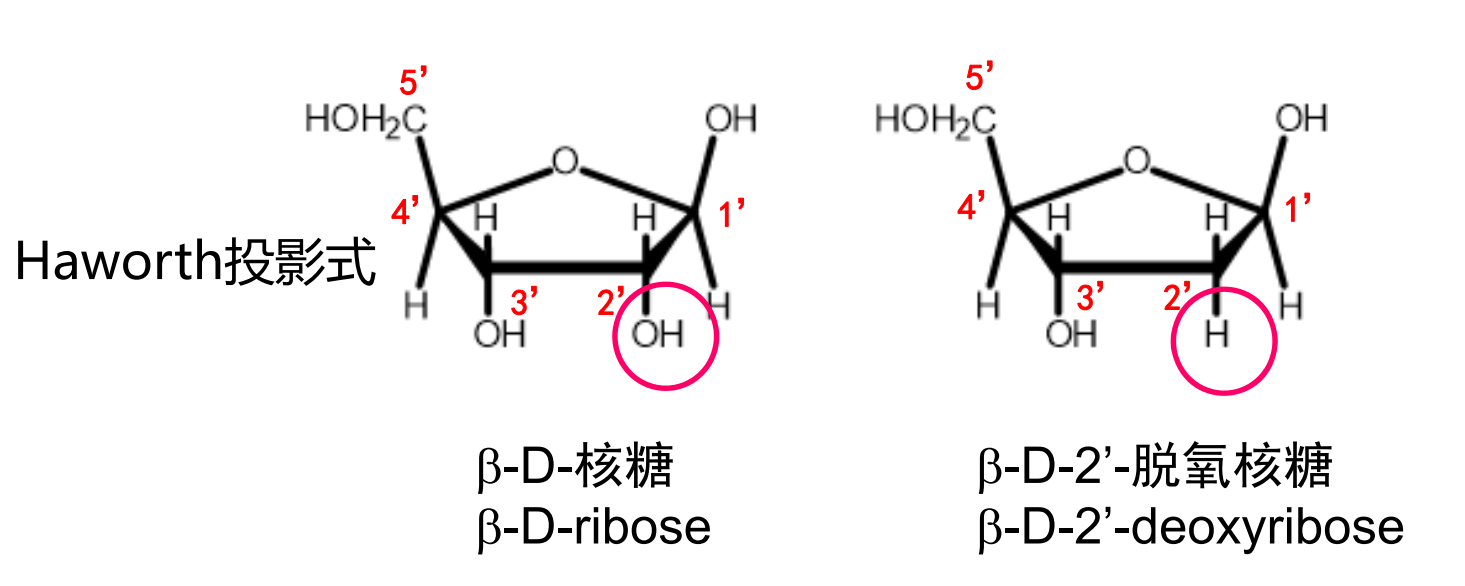
核糖的主要作用是为了连接与支架,它和磷酸一起搭建起了核酸的骨架。
其中脱氧核糖在2号碳上缺少一个羟基。
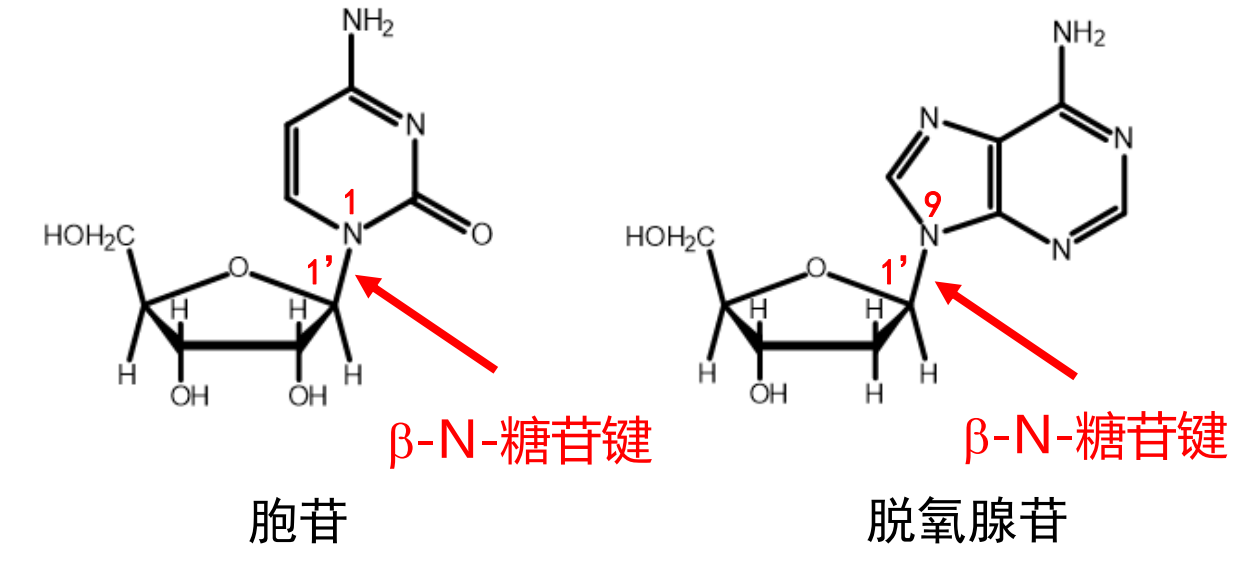
如果我们将碱基和核糖连接起来,就得到了核苷,碱基和核糖之间的化学键叫做\({\beta - N - 糖苷键}\)。
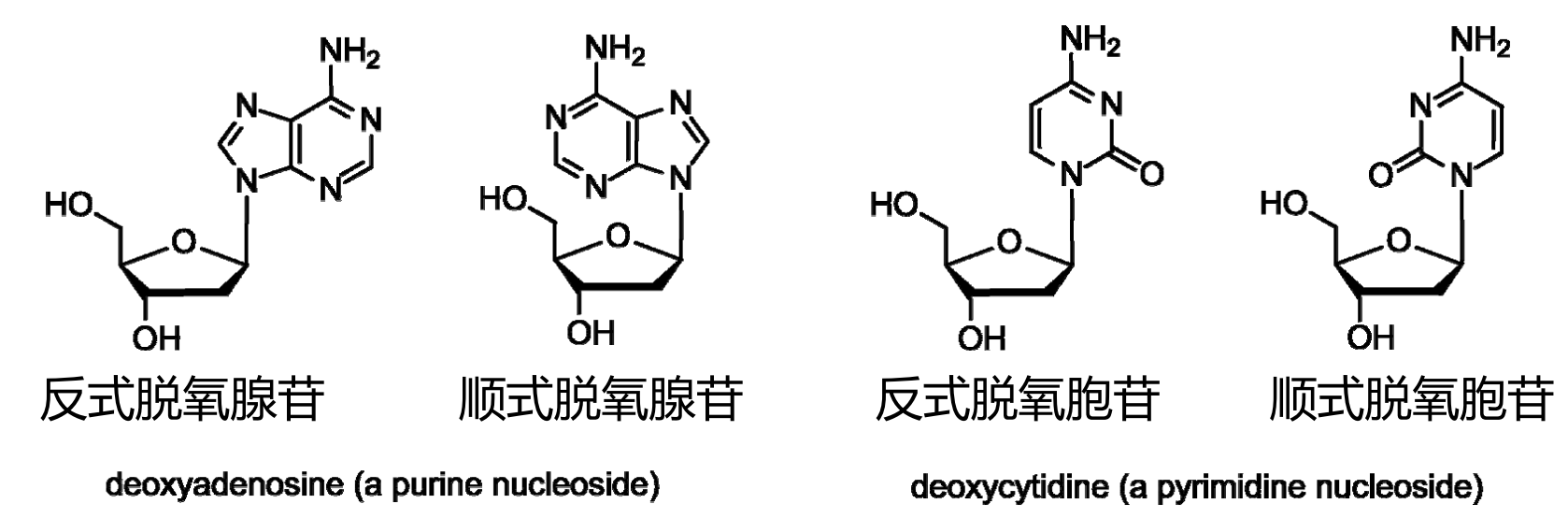
由于空间位阻的影响,碱基和戊糖环就着糖苷键形成反式构象。
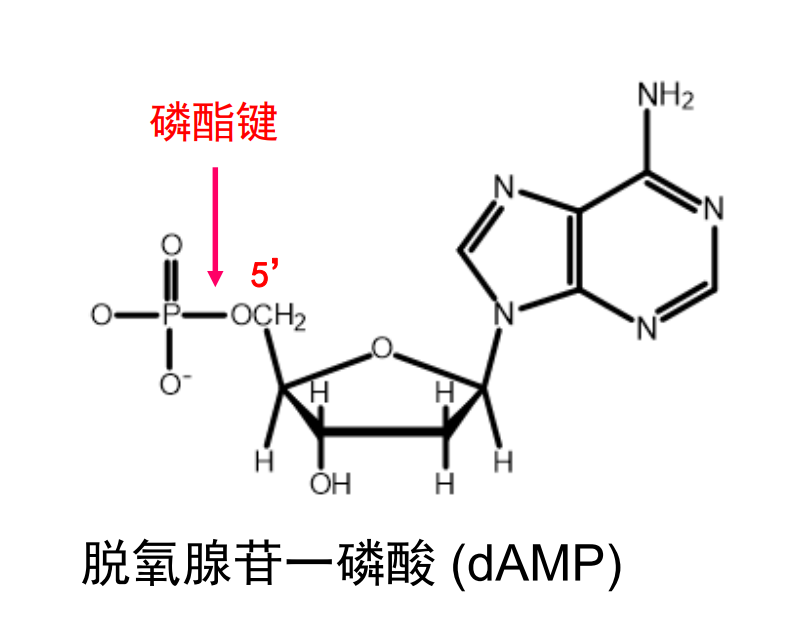
核糖的五号碳上还留有一个高高翘起的羟基,如果拿来一个磷酸基团与其酯化后就能得到核苷酸,磷酸与核糖之间的化学键称之为磷脂键。
仅有一个磷酸基团的核苷酸被称为xxx苷一磷酸(比如脱氧腺苷一磷酸 dAMP),磷酸之间还可以叠叠乐,通过酸酐键相连,形成dNDP,dNTP等等。
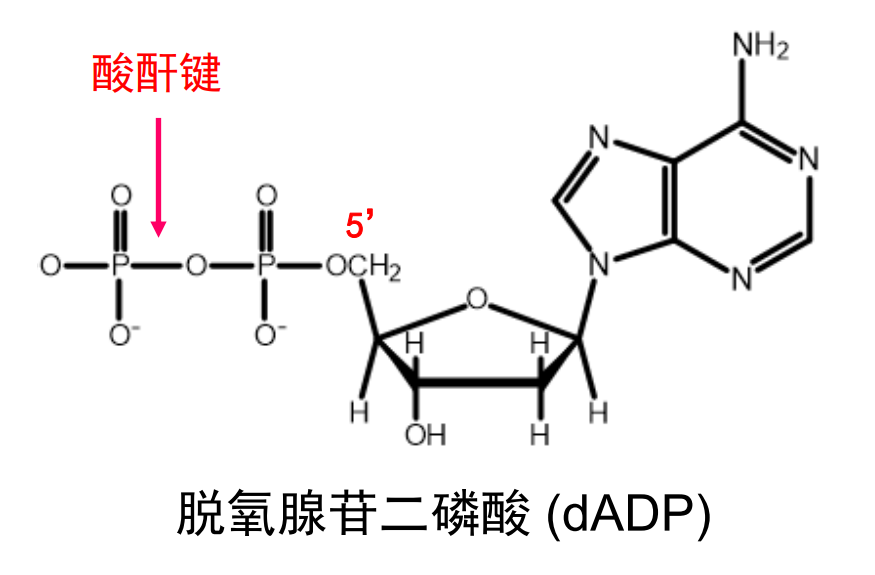
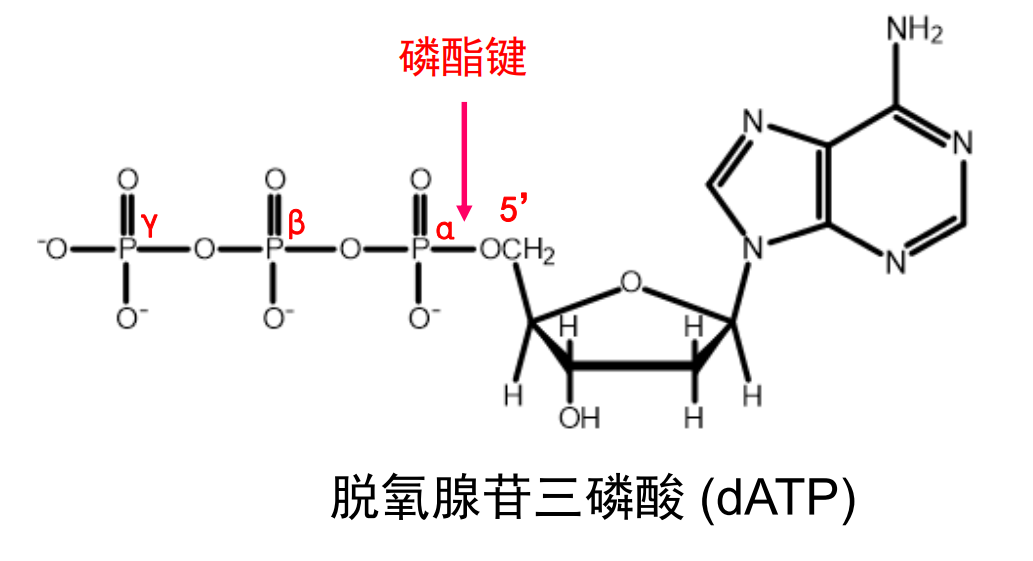
核苷酸的生物学功能
核苷酸本身除了是核酸的基本构成单元以外,还在许多生命活动中充当重要角色,比如作为化学能的载体 (ATP),细胞信号传导分子 (cGMP, cAMP),充当辅酶分子 (NAD+, NADP+, FAD与CoA),肿瘤化疗药物 (6-MP, araC 与 5-FU)。
cAMP/cGMP
这里的"c"的意思是"cyclic",代表"成环",磷酸基团分别与3,5号碳上的羟基形成磷脂键。
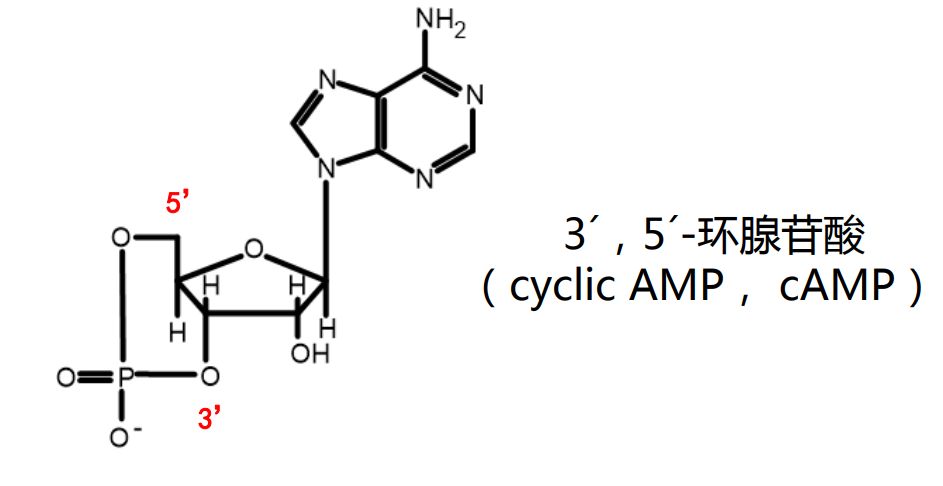
核苷酸组成核酸
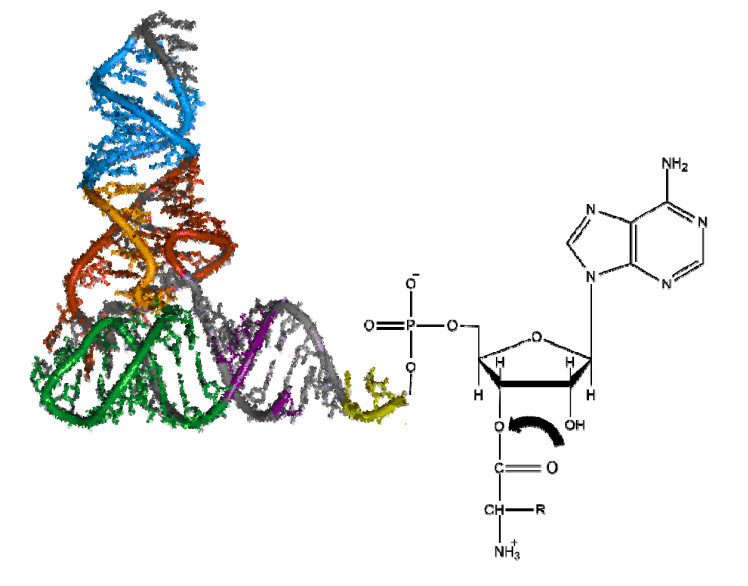
核苷酸之间再次酯化,形成3'-5'磷酸二酯键,相互连接形成DNA/RNA。
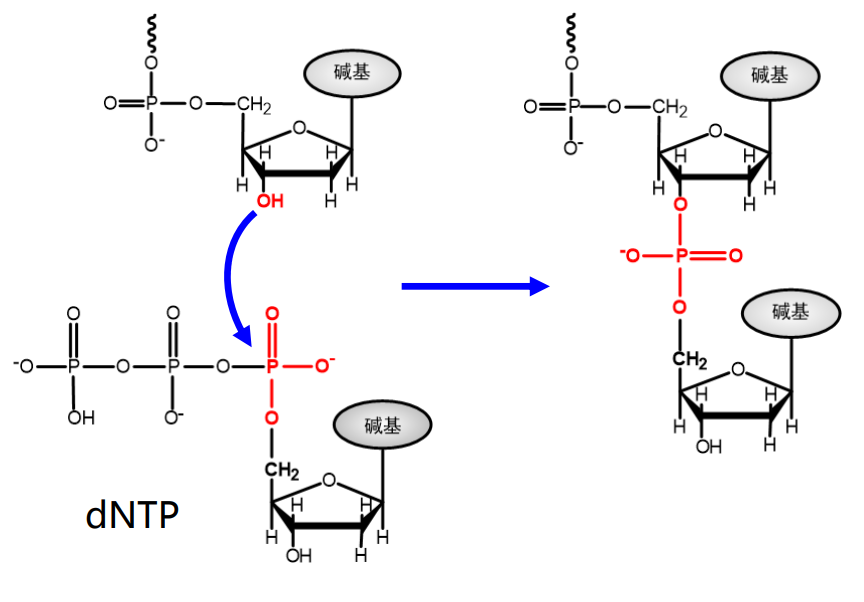
对于多聚核苷酸链来说,其具有方向性,后来者只能从3'-端延长链,也就是说 DNA/RNA 的正方向可以被定义为 5'-3'。
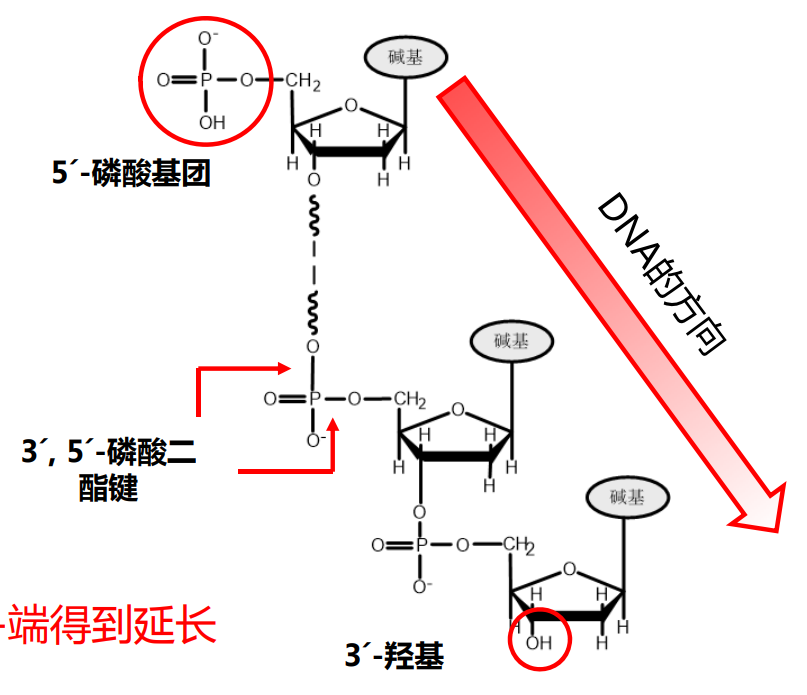
核酸的一级结构
核酸的一节结构与蛋白质的一级结构类似,指的是核苷酸从5'-端到3'-端排列的顺序。
核酸分子的长度用核苷酸数目 (nt) 或者碱基对数目 (bp) 来表示,少于50nt的核酸称为寡核苷酸。
DNA的空间结构与功能
DNA的空间结构 (spatial structure) :构成DNA的所有原子在三维空间的相对位置,包括二级结构与高级结构等等,主要有氢键、离子作用力、疏水作用力和空间位阻效应参与。
DNA二级结构——双螺旋结构
DNA由两条反平行的脱氧核糖核酸链组成,之间由碱基互补配对形成的氢键连接,亲水性骨架将互补碱基对包埋在DNA双螺旋内部,碱基对重叠产生碱基堆积作用,一个螺旋周期包含约10.5bp的碱基对,整个螺旋大多数时候为右手螺旋(Z型 DNA 为左手螺旋)。
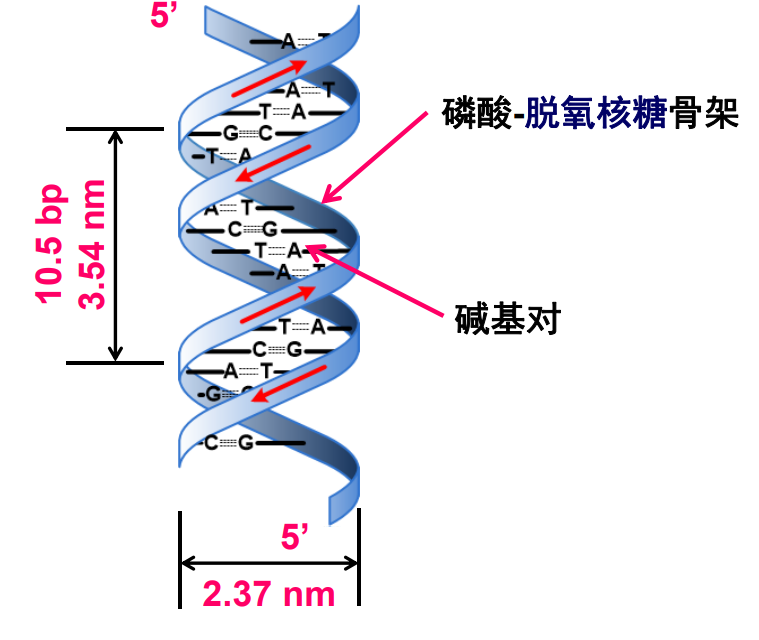
碱基堆积作用 (base stacking)
碱基实际上都有芳香环,相互成对沿着双螺旋骨架叠放,离域电子之间会产生作用,这种作用就是碱基堆积作用,它是稳定DNA双螺旋的重要分子间作用力。
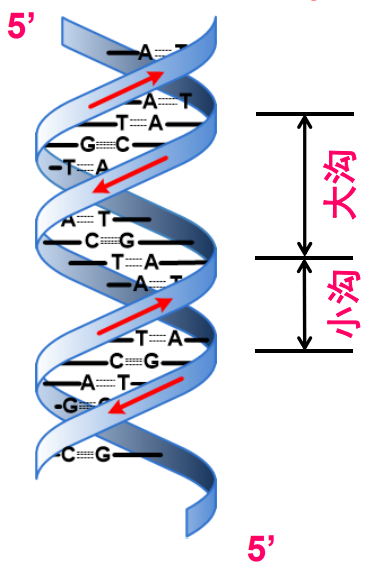
大沟和小沟 (Major groove / Minor groove)
由于碱基的互补配对不是完全的正对,因此一对互补的核苷酸在糖苷键上形成了两侧大小不一的夹角,在双螺旋的结构中(特别是B型)形成了两面大小不一致的沟壑。
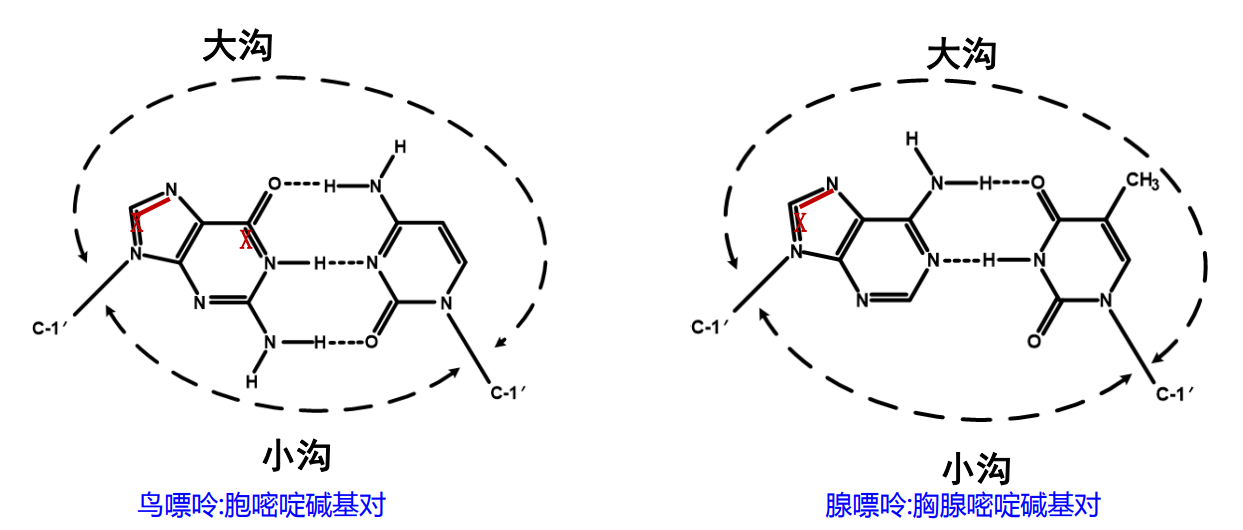
下图则清晰展示了大小沟之间的差距。
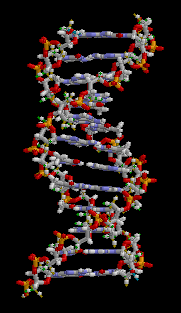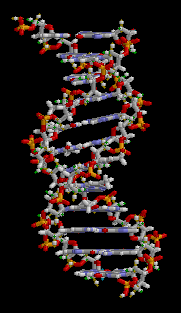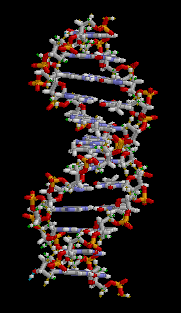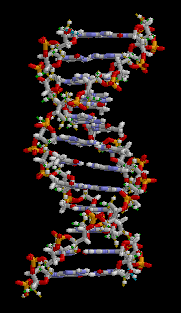
DNA螺旋具有多种不同的形式,常见的有以下三种:
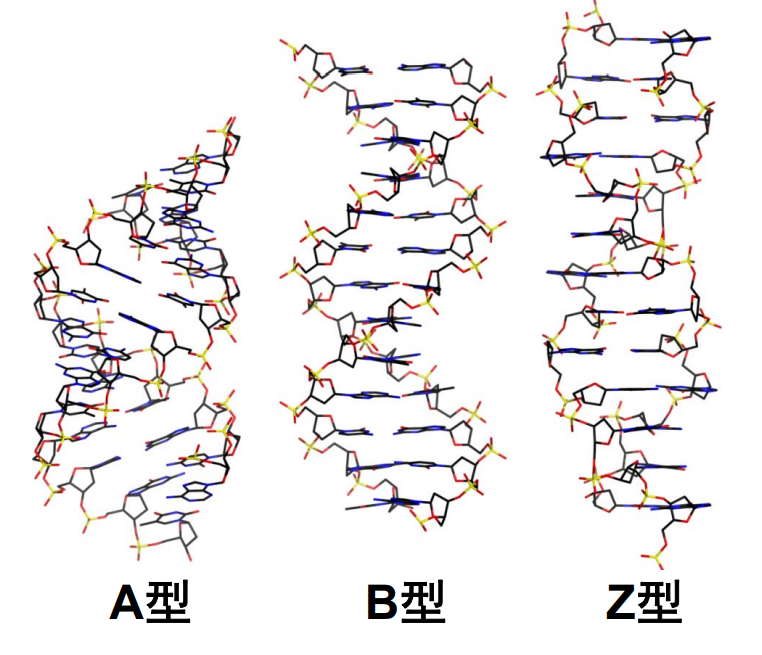
其中 B型就是我们刻板印象里的那种DNA。
而基于氢键互补配对的理论,在GC对和AT对的碱基上仍保有形成氢键的潜力,所以自然而然可以猜想到可能存在的三链螺旋结构。
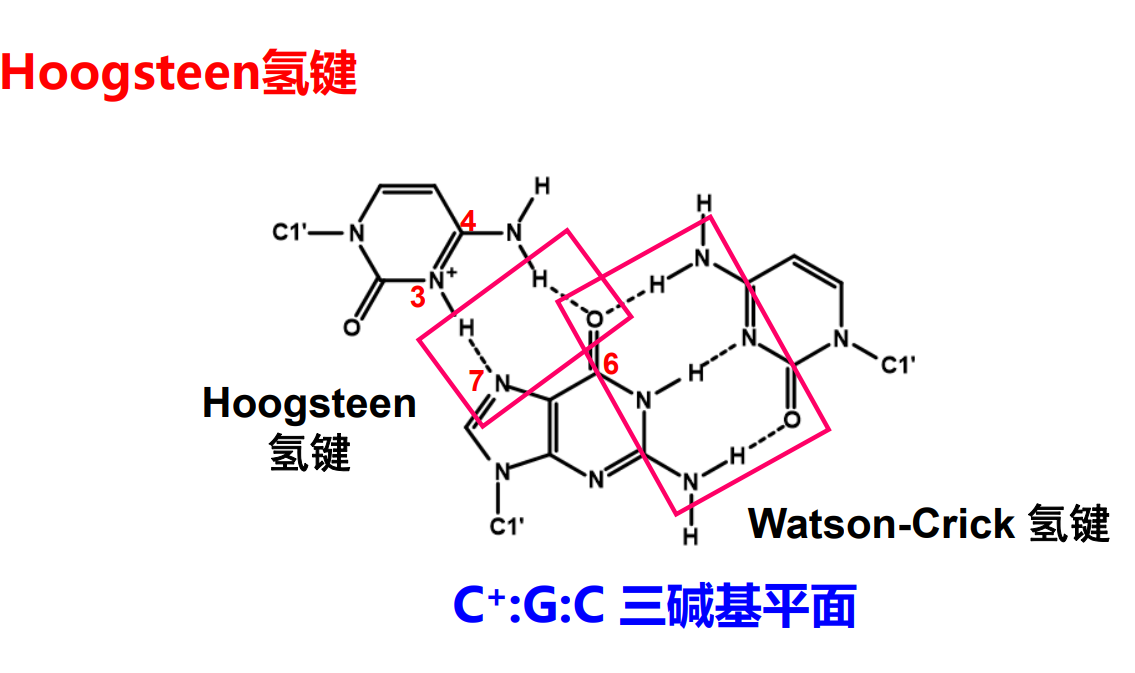
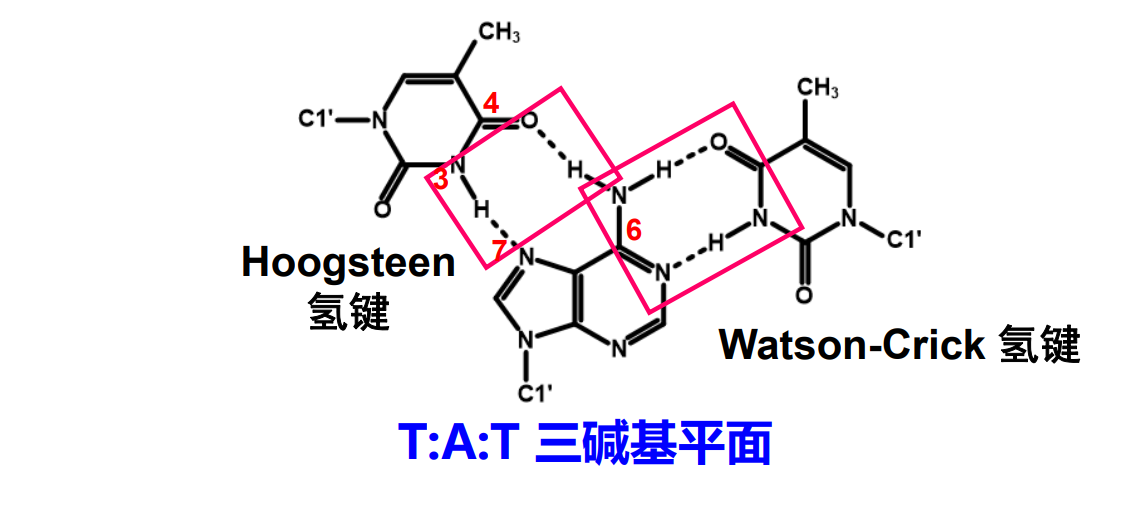
更夸张的,在人的端粒结构上还能出现四链螺旋的结构。
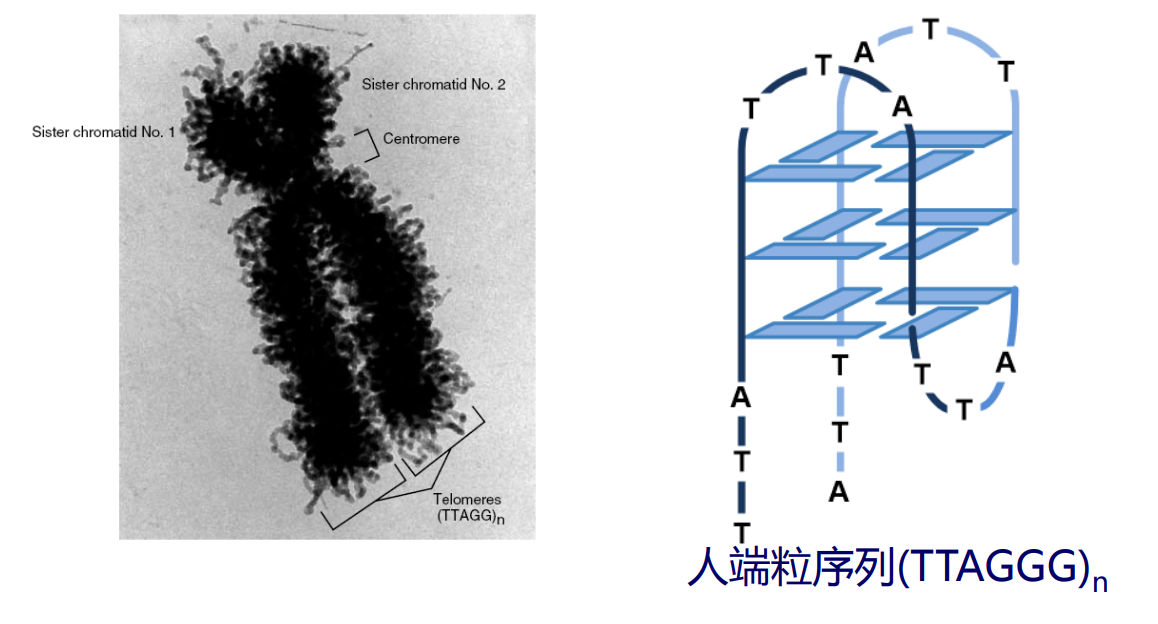
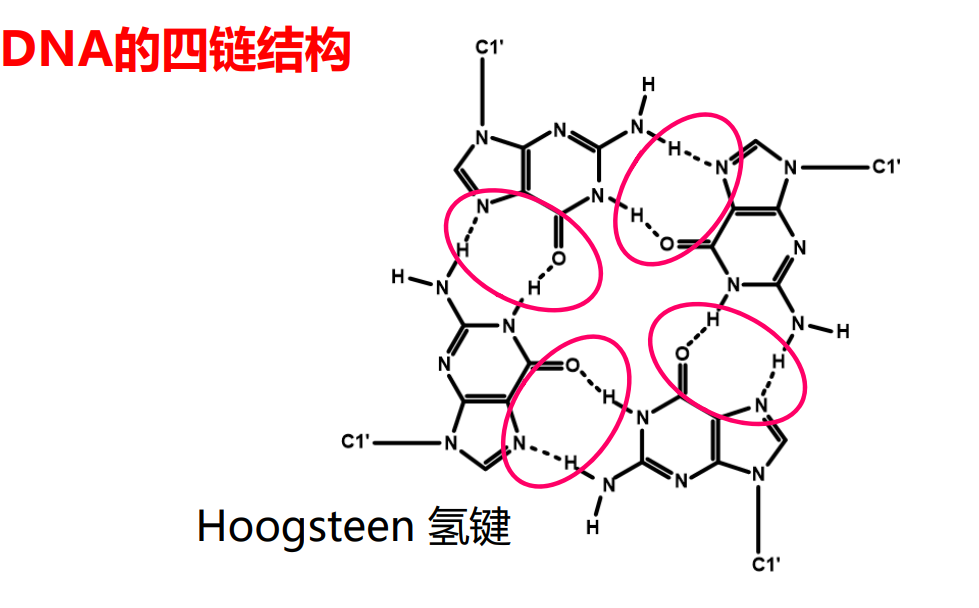
DNA的高级结构
在二级结构基础上继续形成的结构被称为DNA的高级结构。
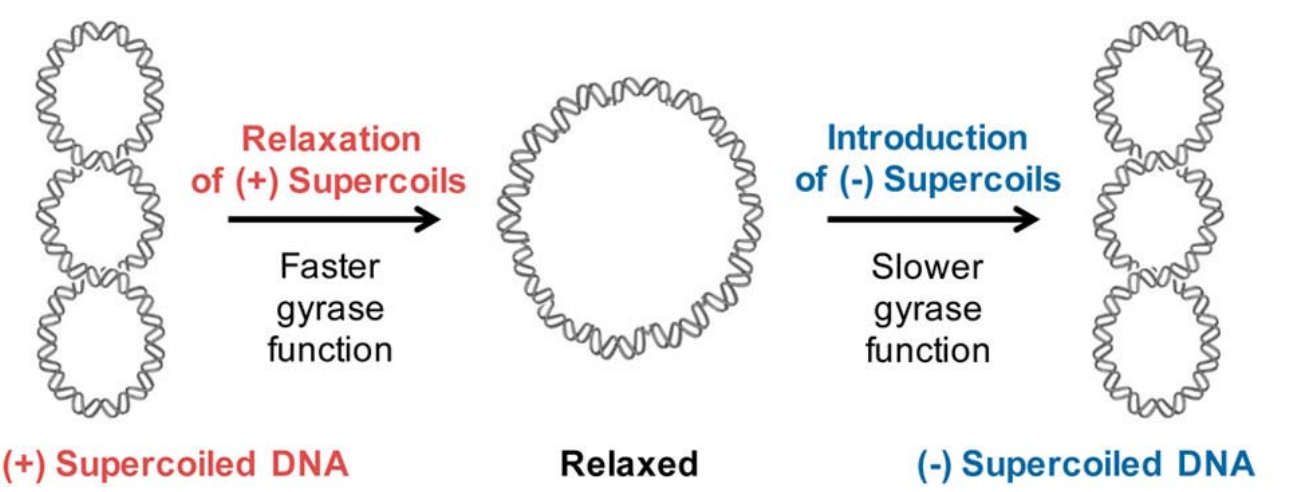
超螺旋
可以想象你将一个环状的橡皮筋向着一个方向拧紧,环状的橡皮筋会自发的开始形成螺旋,这个结构就类似于 DNA 的超螺旋结构(严谨来说,类似于正超螺旋)。
显然,与橡皮筋不同的是,DNA本身的二级结构具有螺旋方向上的取向,如果超螺旋的方向使得双螺旋缠绕更紧,称之为正超螺旋,如果使得双螺旋更加松散,则称之为负超螺旋。
染色质与染色体
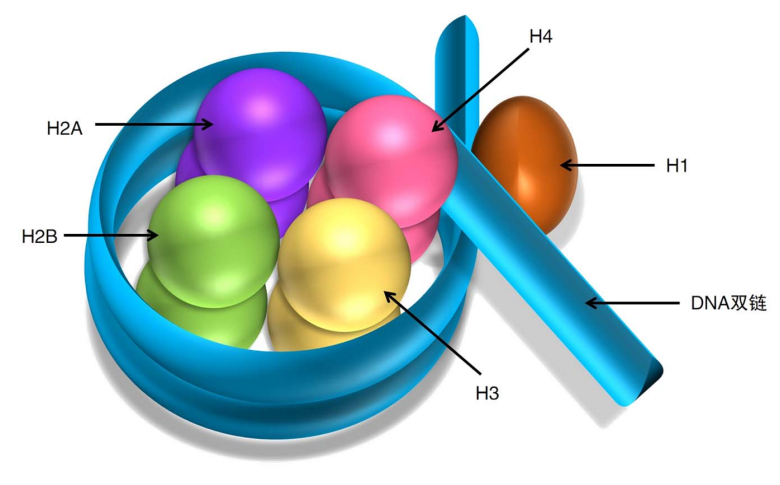
在细胞周期的大部分时间里,DNA以松散的染色质 (chromatin) 形式存在;在细胞分裂期,则形成高度致密的染色体 (chromosome),染色质的基本组成单位是核小体。

核小体的核心是一个组蛋白八聚体,包含有:
- H2A (x2)
- H2B (x2)
- H3 (x2)
- H4 (x2)
还有一个组蛋白H1结合在核小体的进出口处。
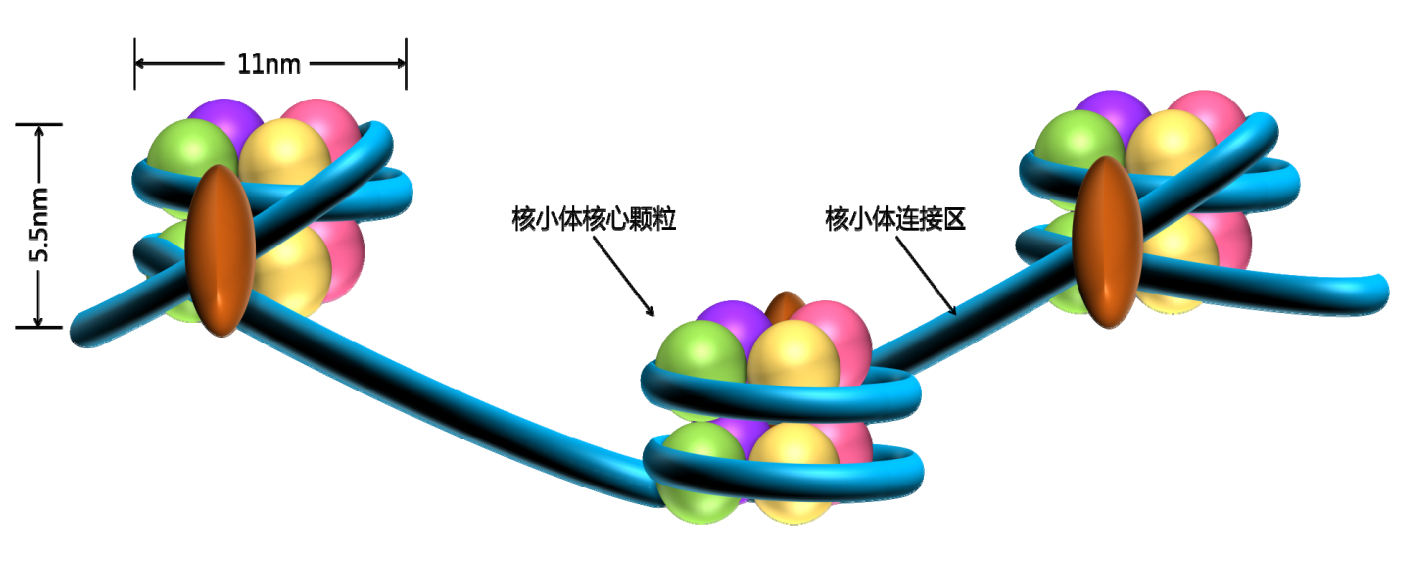
基于核小体再进行进一步的压缩、缠绕,就可以形成更加致密的染色体。
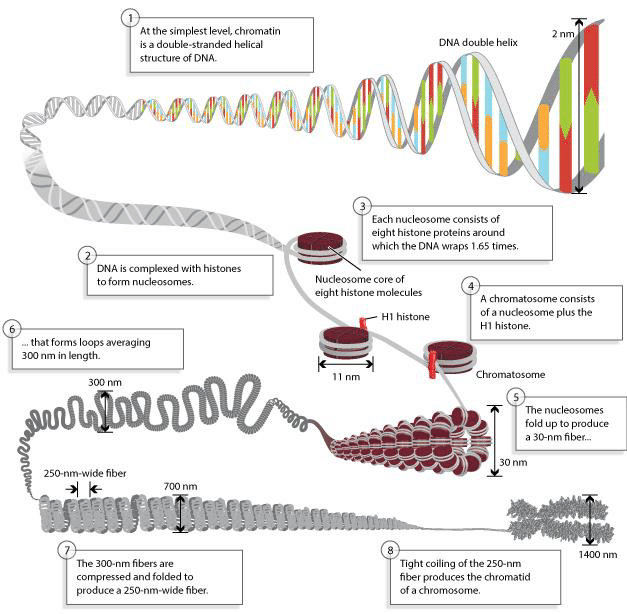
DNA是主要的遗传物质
- 基因 (gene):编码RNA或多肽链的DNA片段,即DNA中一段特定的核苷酸序列。
- 基因组 (genome):包含在该生物的DNA(部分病毒除外)中的全部遗传信息,即一套染色体中的完整的核苷酸序列。
- 病毒颗粒的基因组可以是DNA,也可以是RNA,可以是单链的,也可以是双链的。
RNA的空间结构与功能
mRNA
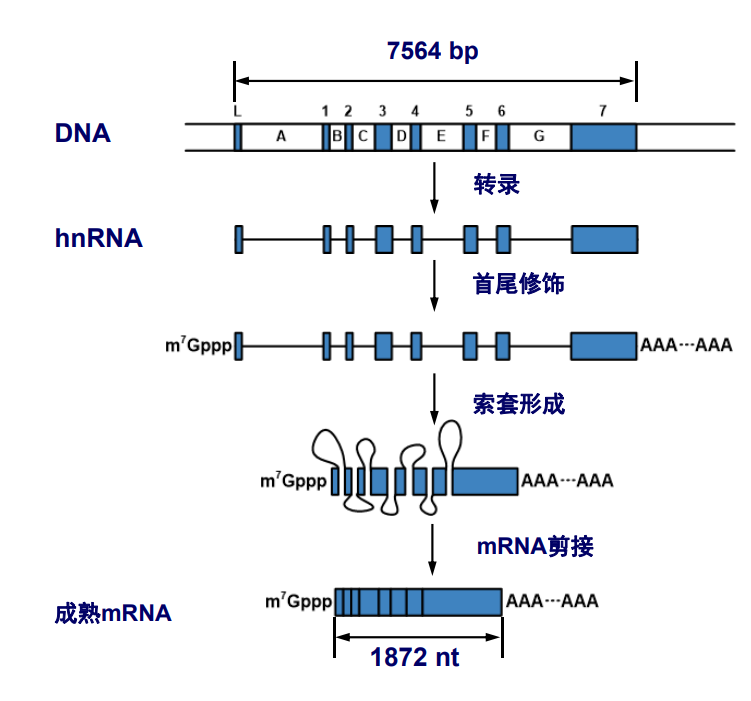
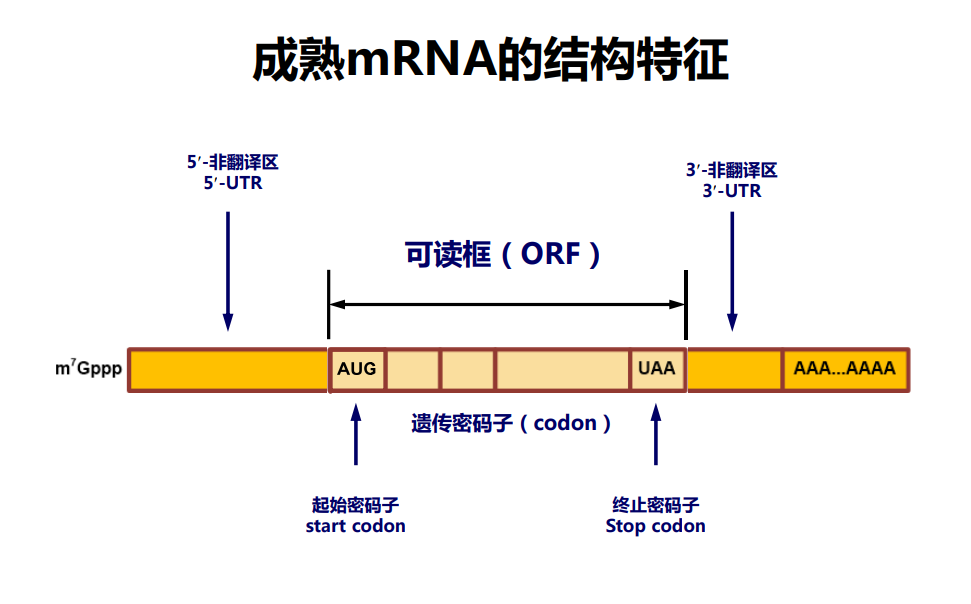
信使RNA,messenger RNA (mRNA),是蛋白质生物合成的模板,它也是生物体内种类最多,丰度最小,差异最大的RNA。由RNA聚合酶II(pol II)合成,细胞核内新生成的mRNA初级产物被称为核不均一RNA(heterogeneous nuclear RNA,简称hnRNA)。
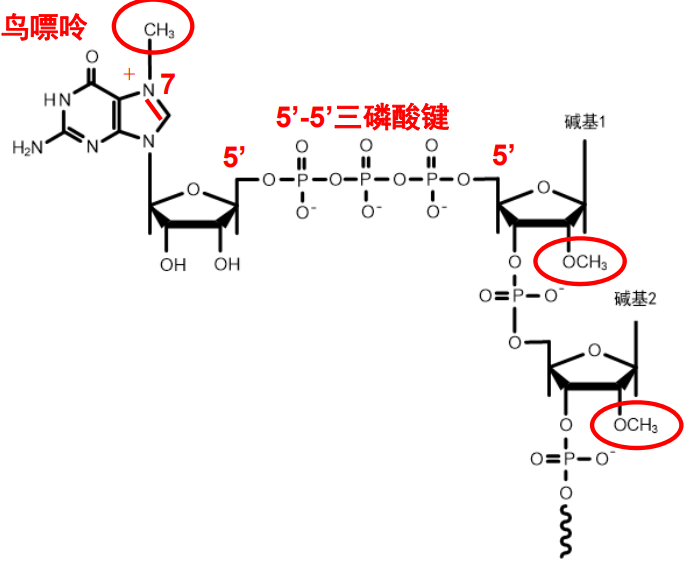
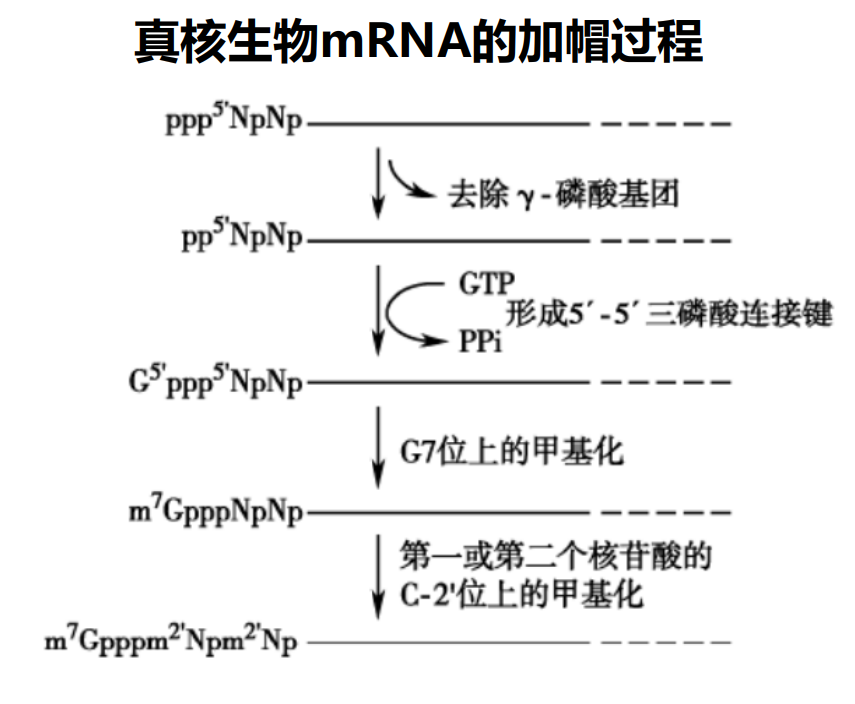
mRNA 5' cap
真核生物的mRNA有5'-端帽结构,具体来说,其在5'-端会有一个通过5'-5'三磷酸键连接的7-甲基鸟嘌呤核苷酸,同时第一个和第二个核苷酸的二号碳上的羟基亦可以发生甲基化,形成一个特殊的“帽子结构”。
5'-帽结构可以与帽结合蛋白 (cap binding protein,CBP) 结合,有助于维持mRNA 的稳定性,协同mRNA 从细胞核向细胞质的转运,以及在蛋白质生物合成中促进核糖体和翻译起始因子的结合。
poly(A)-tail
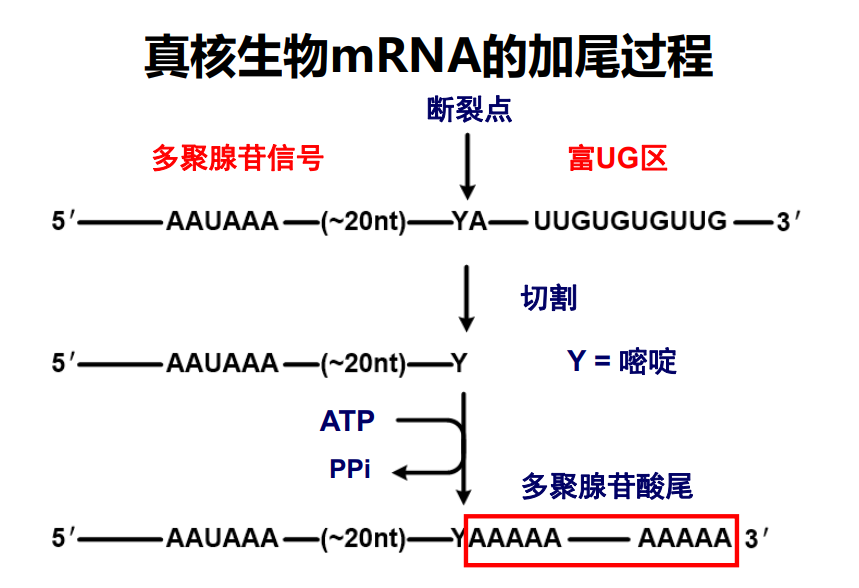
真核生物mRNA的3'-端有一段长度约为80-250腺苷酸,称为多聚腺苷酸尾,可以与poly(A)-结合蛋白 (PABP)结合,与CABP一起同负责mRNA从细胞核向细胞质的转运、维持mRNA的稳定性以及翻译起始的调控。
hnRNA
hnRNA可以视作未成熟的mRNA,是mRNA的前体,其上包含有外显子与内含子。
- 外显子:编码氨基酸的序列。
- 内含子:非编码序列。
hnRNA经过剪接和加工过程,剔除内含子,连接外显子,成为成熟mRNA。
tRNA
转运RNA,transfer RNA(tRNA),作为氨基酸的载体参与多肽链合成,由RNA聚合酶III(pol III)合成,约占RNA总量的15%,长度为74~95 nt,结构非常稳定。
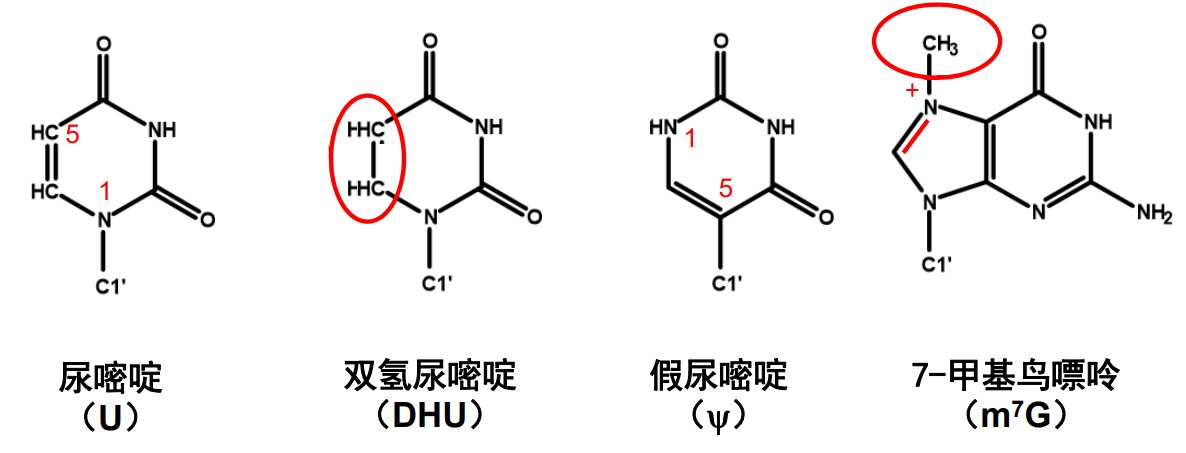
tRNA上含有很多稀有碱基(哇,金色传说!),这些碱基都是在转录后修饰得到的。
tRNA的各个结构拼在一起,在平面上长得像个三叶草,三片叶子分别对应DHU环、反密码子环和TψC环,叶梗对应氨基酸接纳臂。
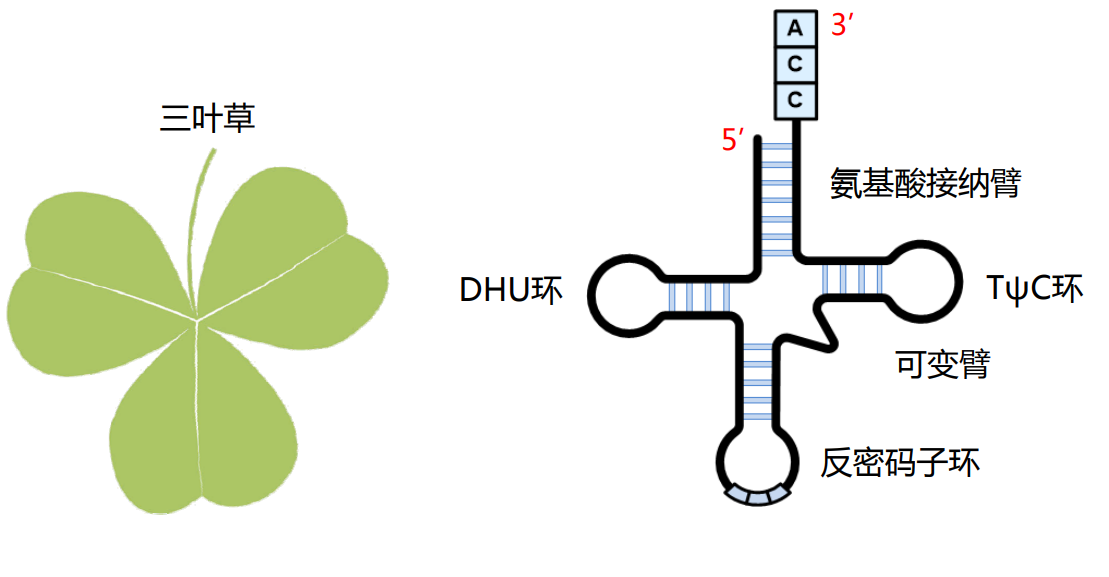
而在空间结构上,其就像一个倒置的“L”。
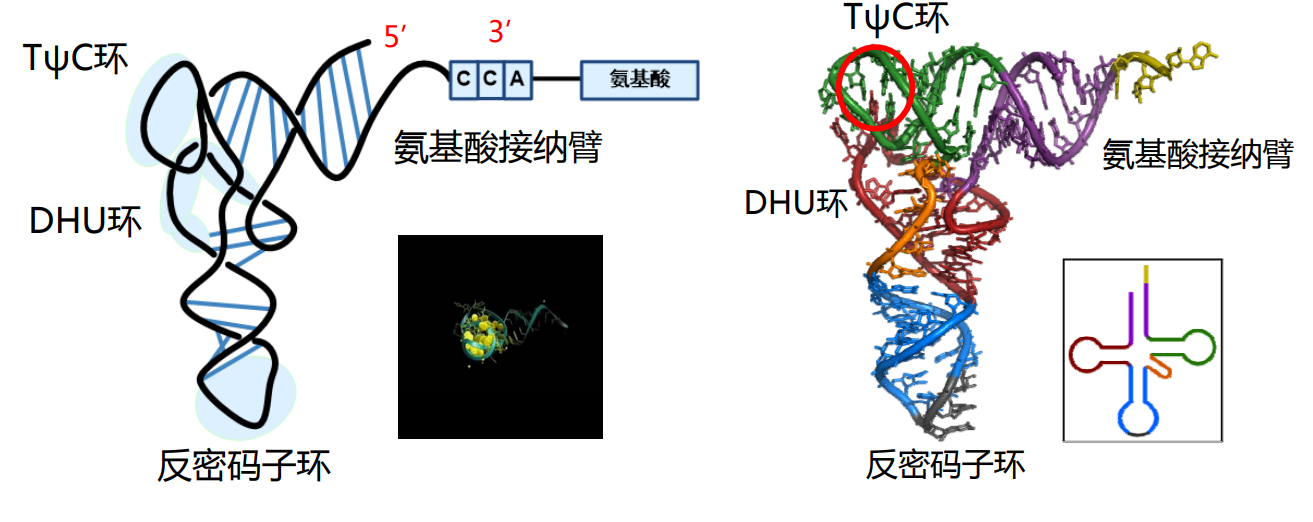
其反密码子环上有三个碱基组成反密码子,其外翻至外侧,方便识别mRNA上的密码子。
tRNA的3'-端负责与氨基酸相连,一种tRNA只能携带一种特定的氨基酸,但是一种氨基酸可以被多种tRNA携带(密码子简并性)。连接在tRNA上的氨基酸才可以用来进行蛋白质的生物合成。
rRNA
核糖体RNA,ribosomal RNA (rRNA),与核糖体蛋白共同组成了核糖体 (ribosome),由RNA聚合酶I和III合成,在原核内有三种,在真核内有四种,是细胞内含量最多的RNA(约占80%),具有稳定的结构和保守的核苷酸序列。
原核和真核的核糖体生物分子组成有所区别:
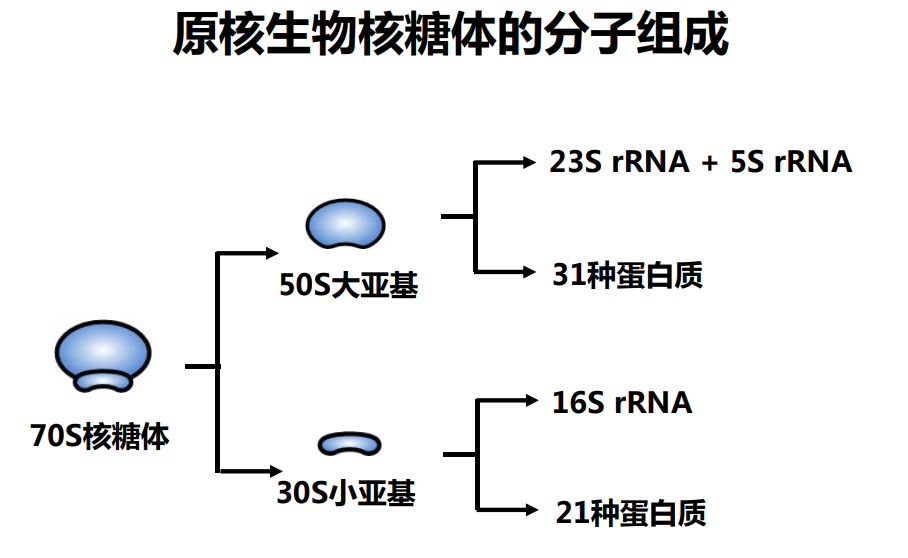
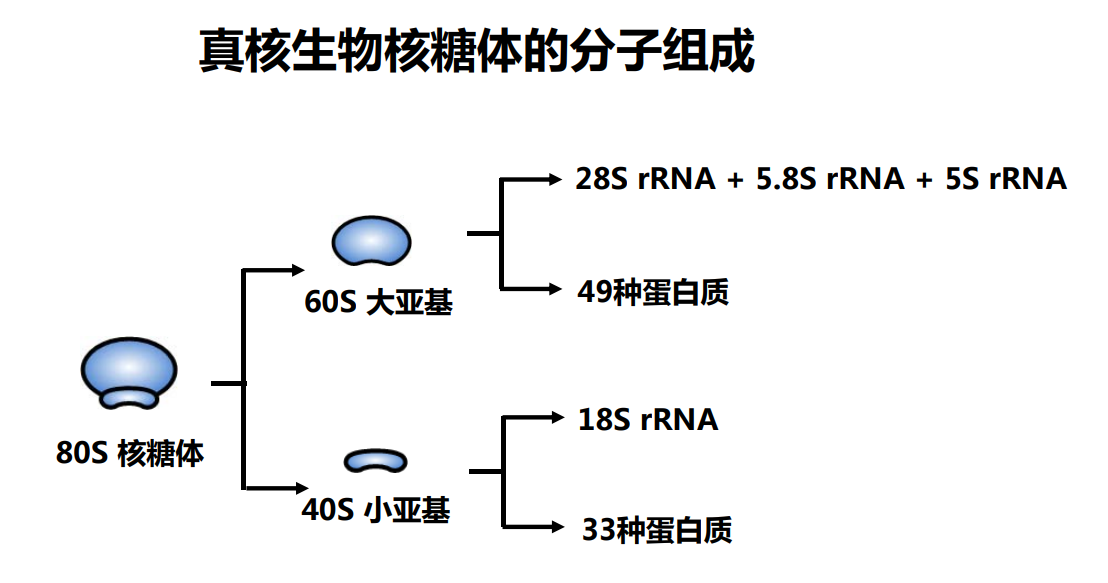
核糖体由大小亚基组成:
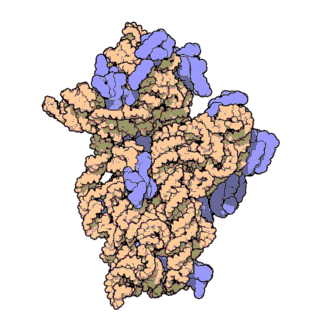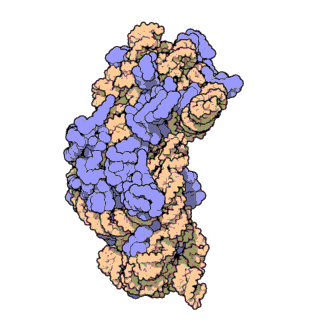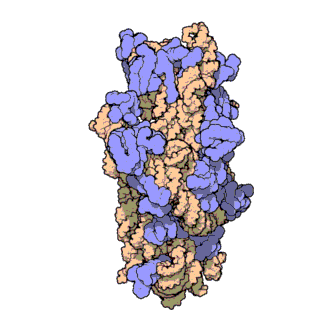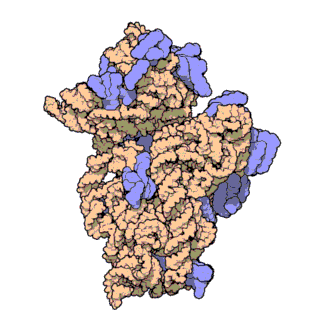
原核生物中核糖体合成蛋白质的过程为:
- 氨酰-tRNA进位A
- 肽链延长(肽链从P位转移到A位)
- 核糖体转位(前进一个密码子)
- tRNA退位(前一个tRNA转移至E位上后退位)

ribozyme
核酶 ribozyme,也称为催化小RNA,是一类具有催化功能的小RNA。
其它组成性非编码RNA
- 核仁小RNA(small nucleolar RNA,简称snoRNA): snoRNA定位 于核仁;参与rRNA的加工,如rRNA的核糖2´羟基的甲基化和假尿嘧啶 化修饰。
- 核小RNA(small nuclear RNA,简称snRNA):参与真核细胞mRNA 的成熟过程,如识别hnRNA上的外显子和内含子的接点,切除内含子。
- 胞质小RNA(small cytoplasmic RNA,简称scRNA): scRNA存在 细胞质中;与蛋白质结合形成复合体后发挥生物学功能,如SRP-RNA与 六种蛋白质共同形成信号识别颗粒,引导含有信号肽的蛋白质进入内质 网进行合成。
非编码小RNA
非编码小RNA,sncRNA,长度小于200碱基,主要包括微RNA (miRNA),干扰小RNA (siRNA),piwi-interacting RNA (piRNA)。
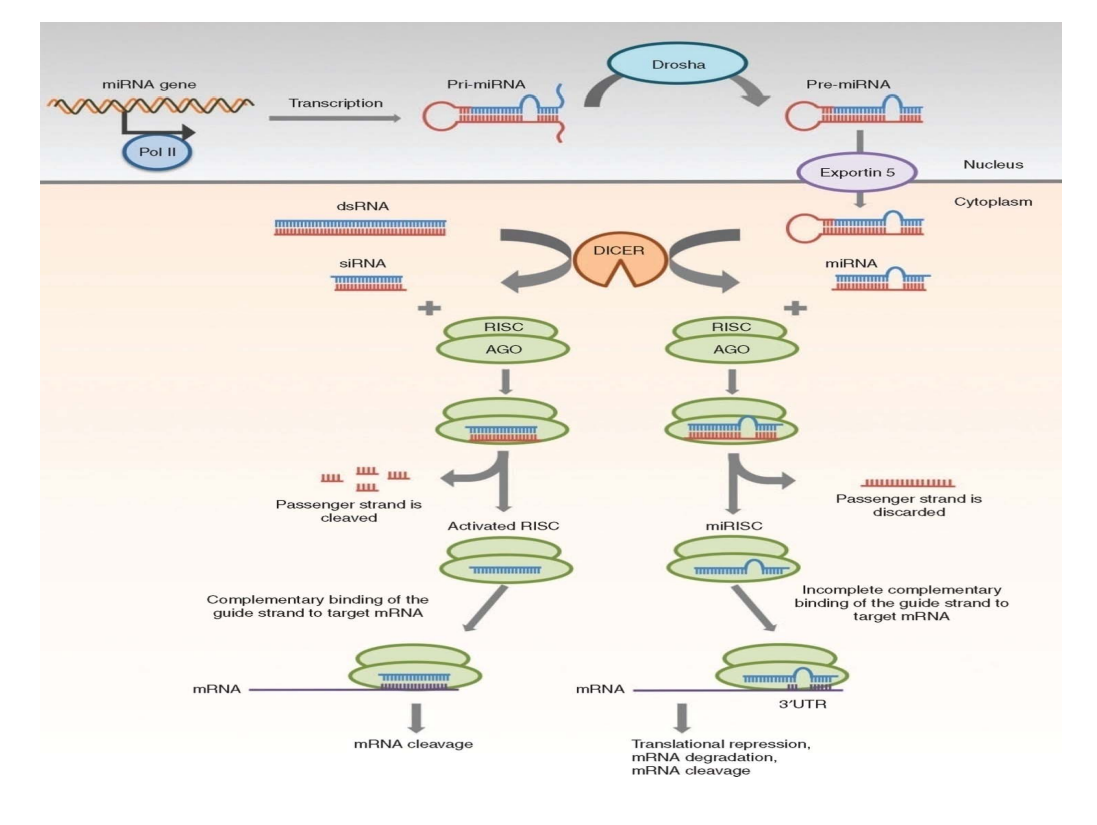
piRNA
- piRNA是哺乳动物生殖细胞中分离得到的~30nt的小RNA,5′端具有强烈的尿嘧啶倾向性(约86%)。
- piRNA与PIWI蛋白家族成员相结合才能发挥调控作用。
- piRNA在染色体上的分布不均匀,piRNA的表达具有组织特异性,调控着生殖细胞和干细胞的生长发育。
长非编码RNA
长非编码RNA,lncRNA,长度为200-100,000个核苷酸,具有poly(A)尾,但不存在ORF,来源于蛋白质编码基因、假基因或编码基因间的DNA序列,由RNA聚合酶Ⅱ转录,经剪切加工后生成,具有强烈的组织特异性与时空特异性。
lncRNA有以下几点功能:
- 结合在编码基因启动子区,干扰下游基因的表达;
- 与编码基因的转录本形成互补双链,形成不同的剪切形式;
- 与编码基因的转录本形成互补双链,产生内源性siRNA;
- 与特定蛋白质结合,调节相应蛋白的活性;
- 结合到特定蛋白质上,改变该蛋白质的细胞定位;
- 介导染色质重构,影响下游基因的表达。
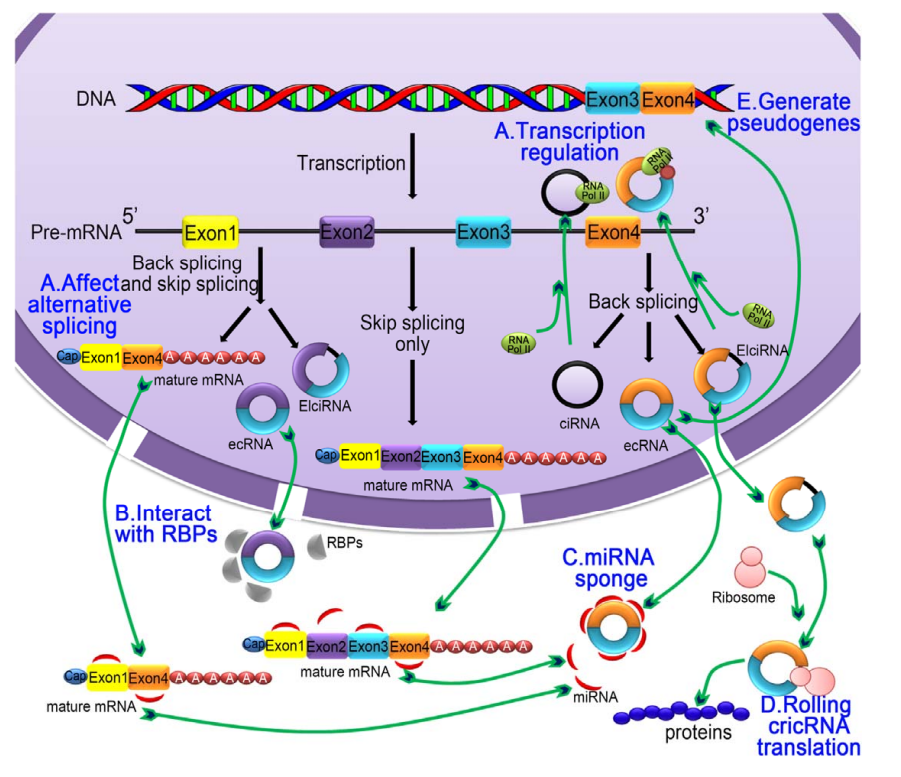
环状RNA
环状RNA不易降解,在细胞中起到miRNA海绵(miRNAsponge)的作用.
核酸的理化性质
紫外吸收
同蛋白质类似,核酸也有强烈的紫外吸收特征(芳环:你好)。
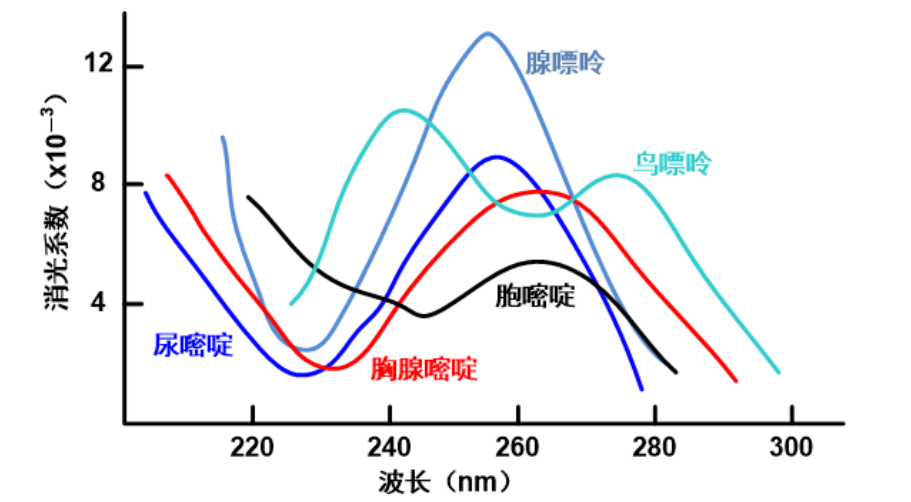
可以看到主要在260nm左右有强烈的紫外吸收特征。
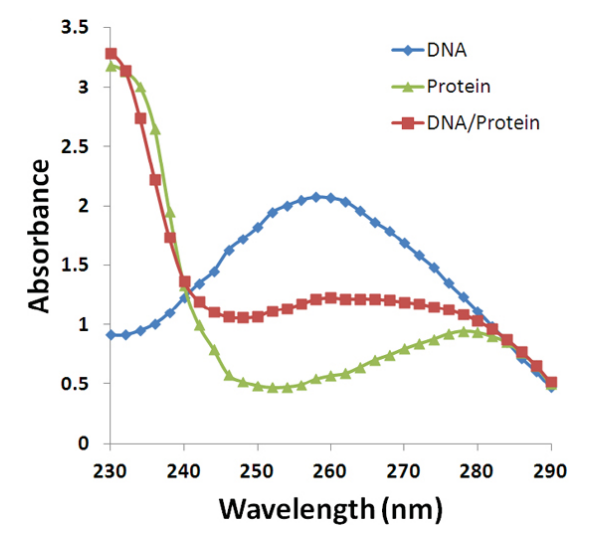
由于蛋白质主要在280nm有较强的吸收峰,所以我们可以通过计算\({\rm{OD_{260}/OD_{280}}}\)的比值来测定样品内核酸与蛋白质的纯度如何。(由于尿嘧啶对260nm的紫外吸收强于胞嘧啶,DNA和RNA也可以就此区分)
- 对于确定样品中DNA或RNA的含量,\({\rm{OD_{260} = 1}}\)等效于
- \(50\mu g/ml\) 双链DNA
- \(40\mu g/ml\) 单链DNA或RNA
- \(20\mu g/ml\) 寡核苷酸
- 对于确定样品中DNA或RNA的纯度
- 纯 DNA: \({\rm{OD_{260}/OD_{280} = 1.8}}\)
- 纯 RNA: \({\rm{OD_{260}/OD_{280} = 2.0}}\)
- 纯蛋白: \({\rm{OD_{260}/OD_{280} = 0.6}}\)
DNA变性
类似于蛋白质,在某些理化因素作用下, 双链DNA(double stranded DNA, 简称dsDNA)解离成为两条单链DNA(single stranded DNA, 简称ssDNA)的过程被称为DNA的变性。
值得注意的是,变性破坏了DNA的空间结构,但是没有改变DNA的序列。
常见的DNA变性因素有::强酸、强碱、加热、变性剂(尿素、甲酰胺)。
DNA的变性存在多个不同的阶段,分别对应不同强度的变性条件。
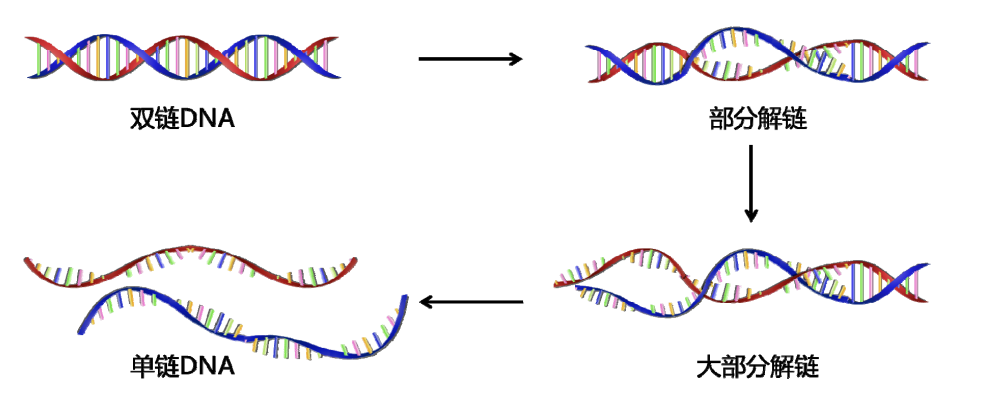
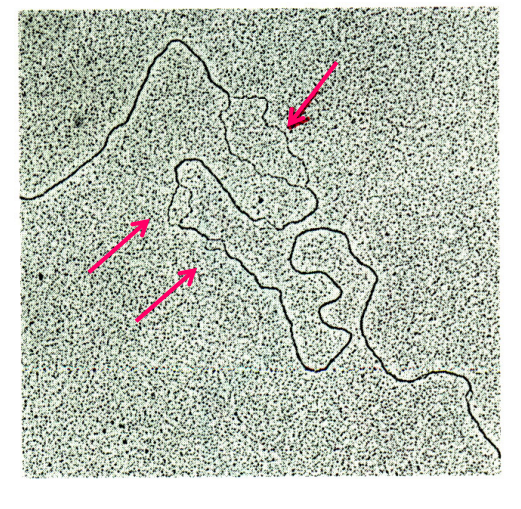
在电镜下可以清晰看到相应的解链片段。
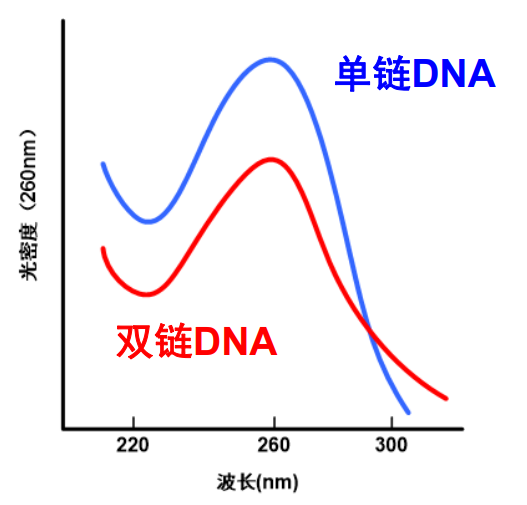
DNA变性后其物理性质也会发生变化,比如DNA变性的增色效应。
DNA变性的增色效应
在DNA变性过程中,它在260nm处的OD值会发生增加。这种变化称为增色效应 (hyperchromiceffect)。
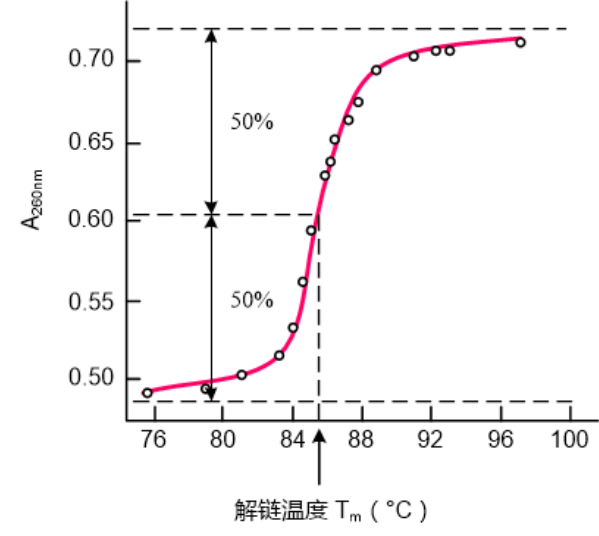
基于这样的性质,我们规定DNA的解链温度 Tm为DNA的解链过程中,紫外光吸收值达到最大变化值的50%时的温度,基于DNA对260nm的吸收程度不同,可以绘制DNA的解链曲线。
:DNA的长度、GC含量、溶液的离子强度均可能影响Tm值。
DNA的复性
逐渐去除变性条件,两条单链DNA会缓慢地形成一条双链DNA,恢复天然的双螺旋结构,这个过程被称为DNA的复性,其中退火特指热变性的DNA经缓慢冷却后的复性。
DNA复性的前提条件是两条链互补,复性后的DNA在260nm的OD水平恢复正常值。
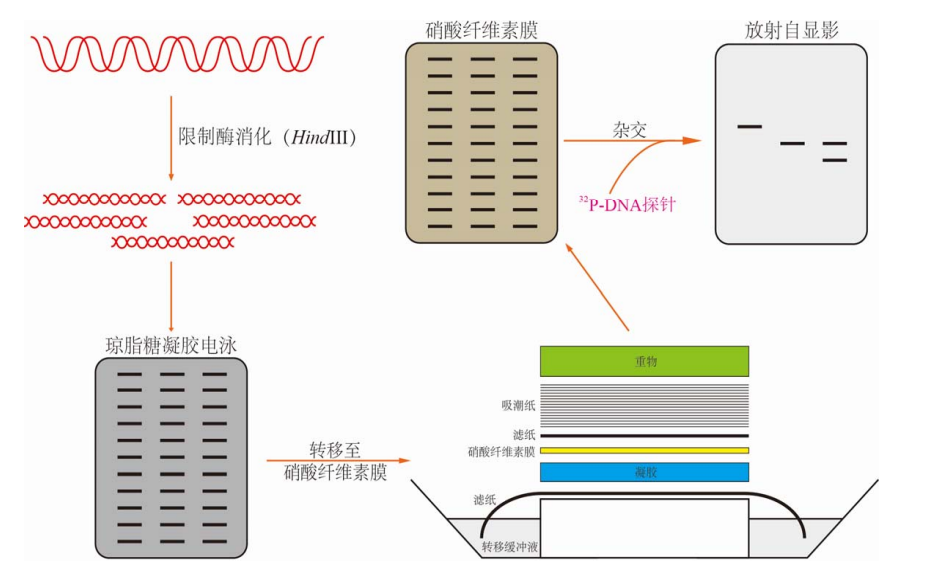
核酸分子杂交
碱基序列互补的两条单链DNA、或两条单链RNA、或一条单链DNA与一条单链RNA形成双链现象。
这个现象基于碱基互补配对的原理,只要一定数量的碱基片段能够匹配上,两条单链就能自发形成部分双链的结构。
用途:
- DNA印迹(Southern blotting)
- RNA印迹(Northern blotting)
- 斑点印迹 (dot blotting)
- 原位杂交 (In situ hybridization)
- 聚合酶链式反应(PCR)
- 基因芯片(DNA microarray)
核酸实验相关
核酸提纯:乙醇沉淀、苯酚-氯仿抽提与试剂盒。
核酸电泳:琼脂糖凝胶电泳。
DNA印迹: