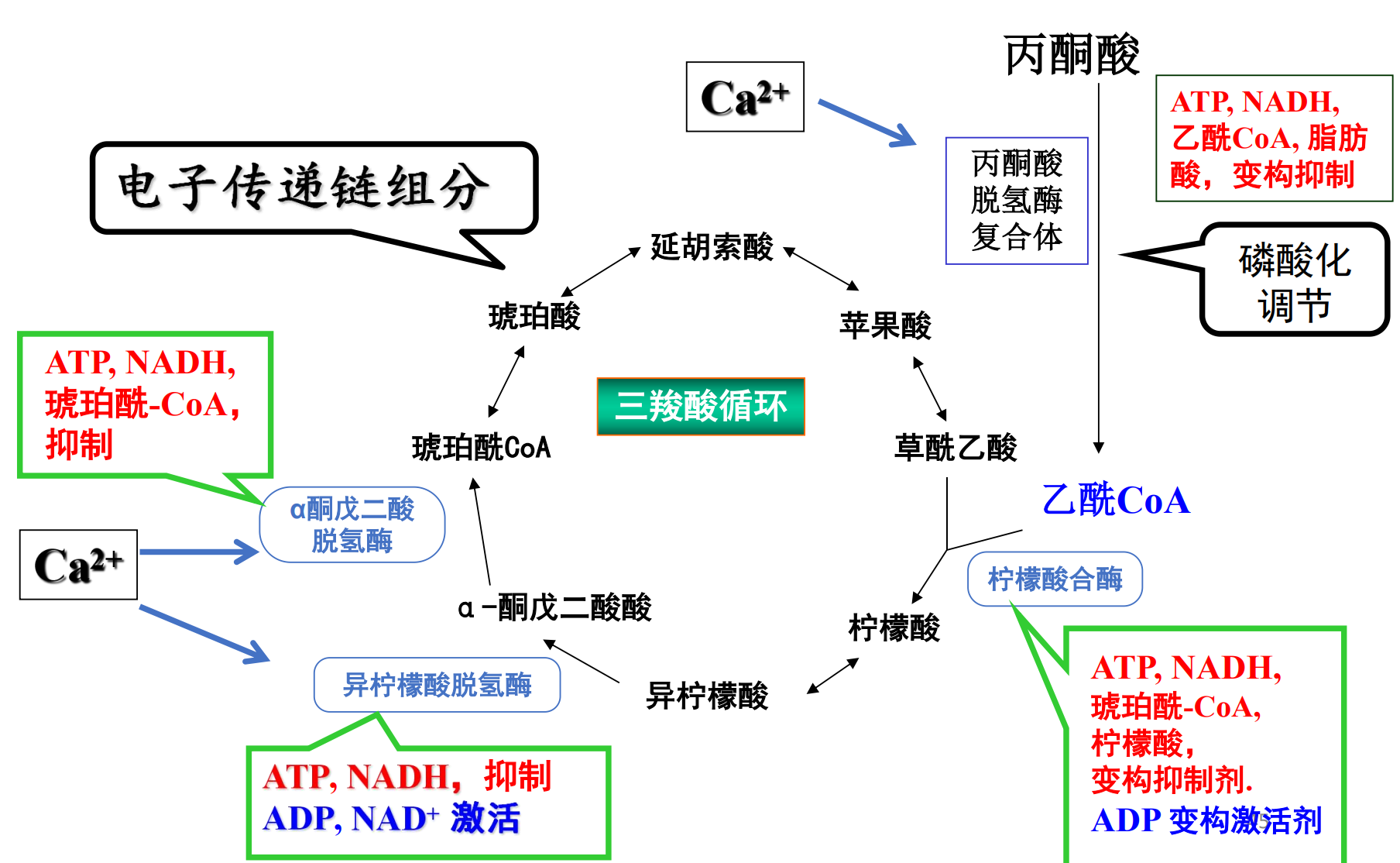
三羧酸循环
三大营养物质——糖、脂肪与部分氨基酸氧化生成乙酰CoA,再通过三羧酸循环氧化乙酰基团并提取出电子,在通过电子传递链最终还原氧气,偶联合成ATP。
三羧酸循环在这个过程中起到承上启下的作用。
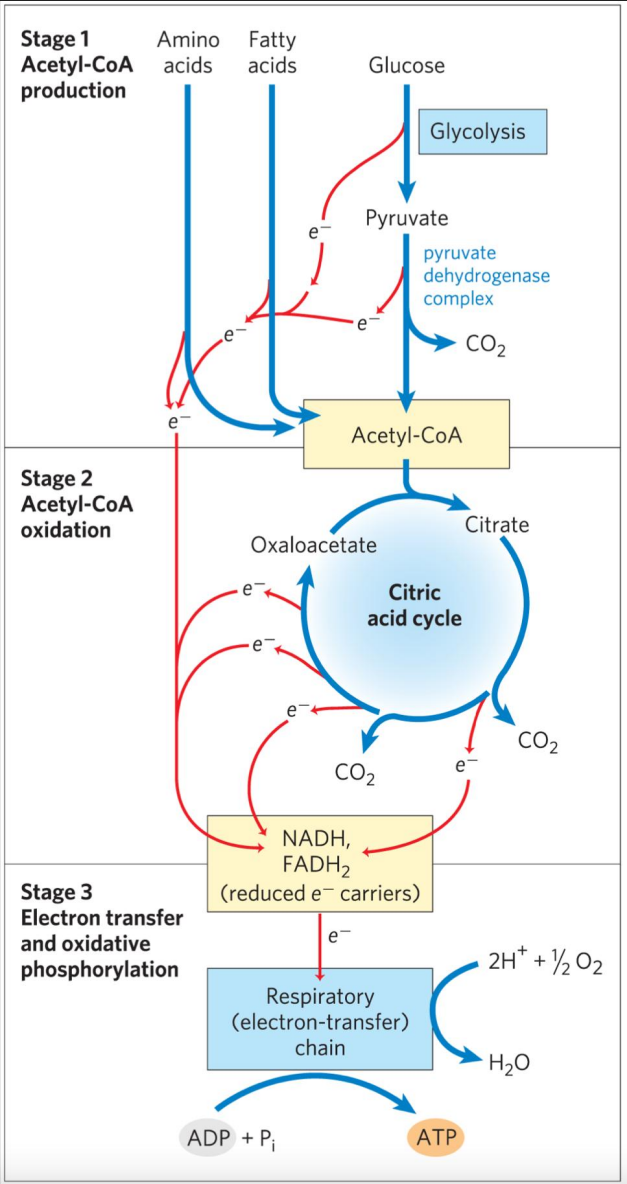
三羧酸循环过程
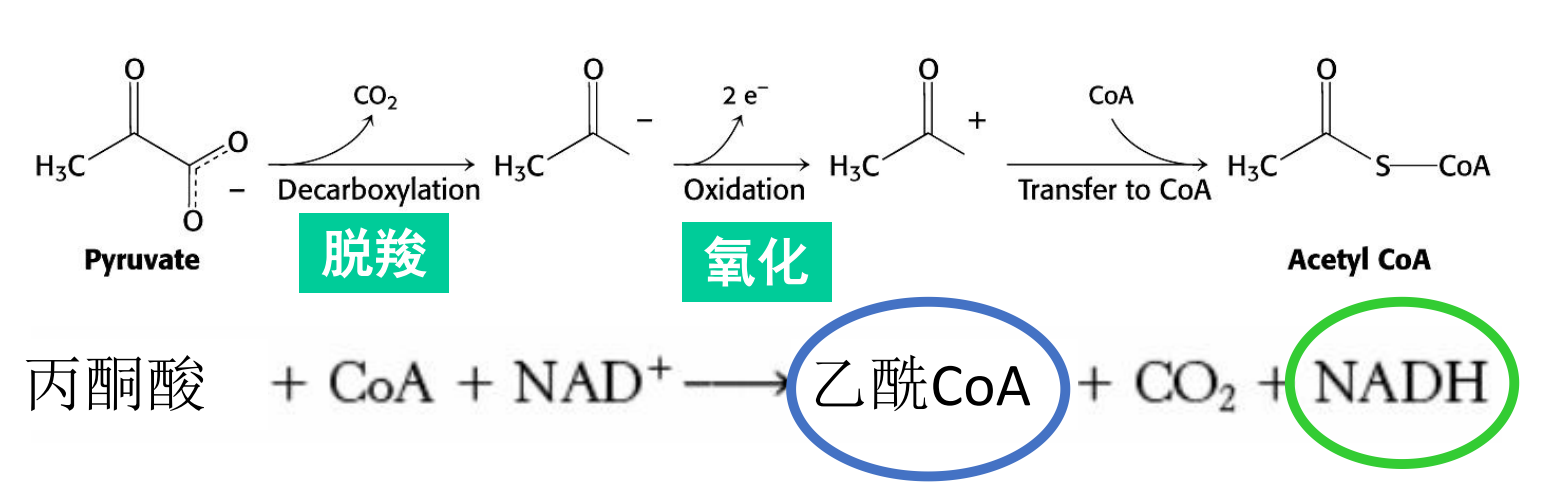
丙酮酸转化为乙酰CoA
丙酮酸脱氢酶复合体催化丙酮酸转化为乙酰CoA,并产生1分子的NADH,这是正式进入三羧酸循环之前的准备阶段。
三羧酸循环概述
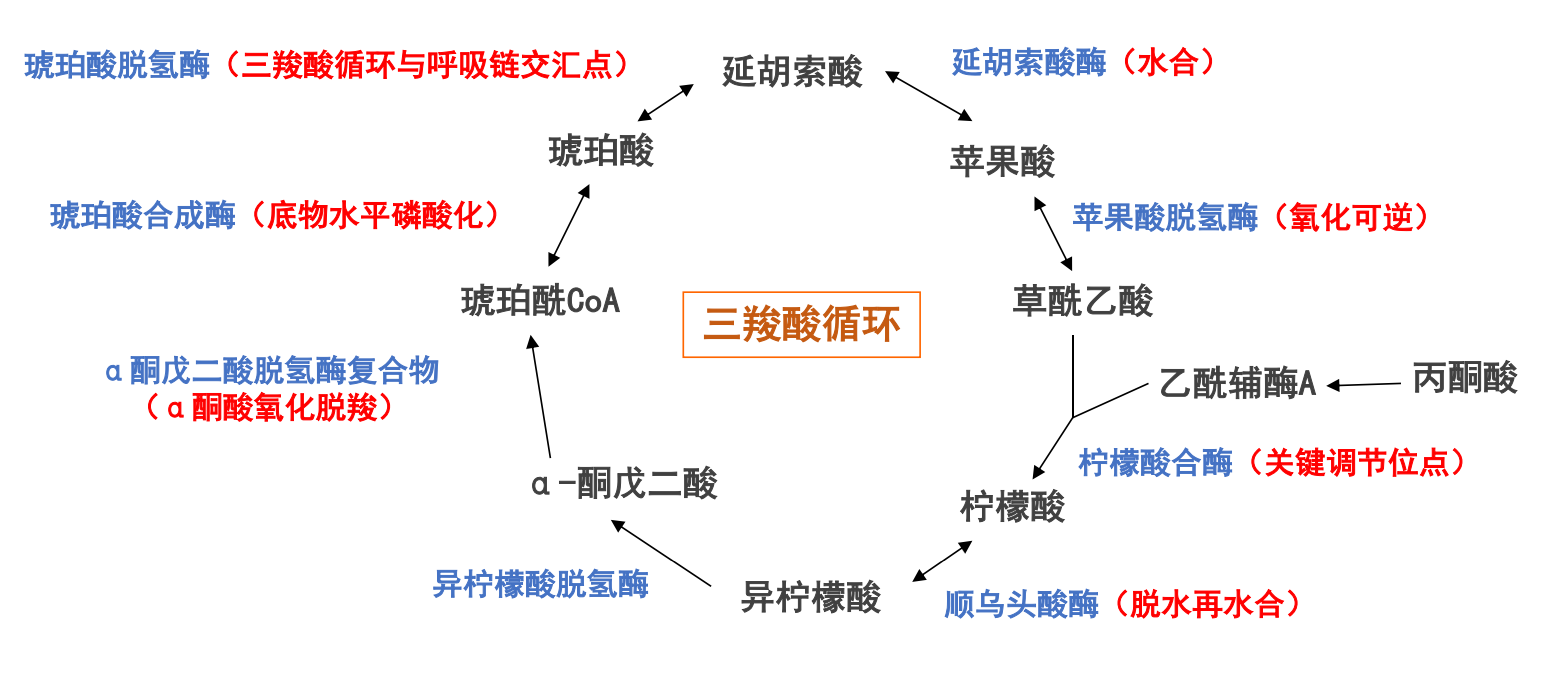
三羧酸发生在线粒体基质,经过8步反应,每次循环氧化一个二碳单位,生成:
- 两分子\(\ce{CO2}\)
- 一分子GTP或ATP
- 乙酰CoA电子转移生成三分子\(\ce{NADH}\)和一分子\(\ce{FADH2}\)
从柠檬酸到草酰乙酸的7步反应等效于乙酸或甲烷的氧化,底物消耗两个碳(碳链长度从6缩减到4),失去8个电子,等效于消耗两分子\(\ce{O2}\):
- 两步脱羧:⍺-酮戊二酸前后
- 两步氧化:琥珀酸脱氢和苹果酸脱氢
- 一/两步异构:柠檬酸-顺乌头酸-异柠檬酸
- 一步水合:延胡索酸-苹果酸
- 一步底物水平磷酸化:琥珀酰-CoA
Note
三羧酸循环有很多名称,包括三羧酸循环 (TCA循环)、柠檬酸循环 ( Citric acid cycle)与Kerbs循环。
三羧酸循环步骤
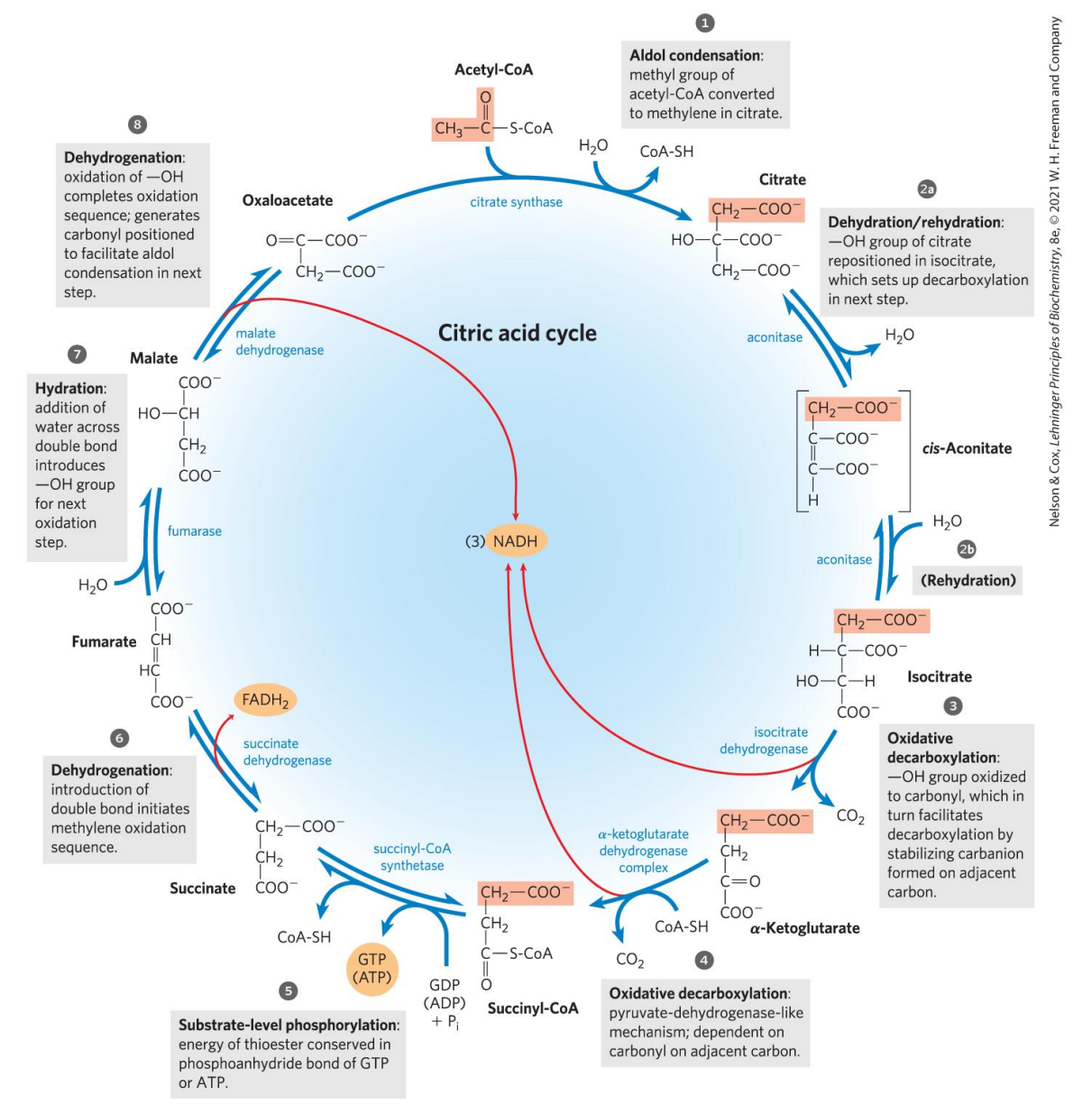
第一步 柠檬酸
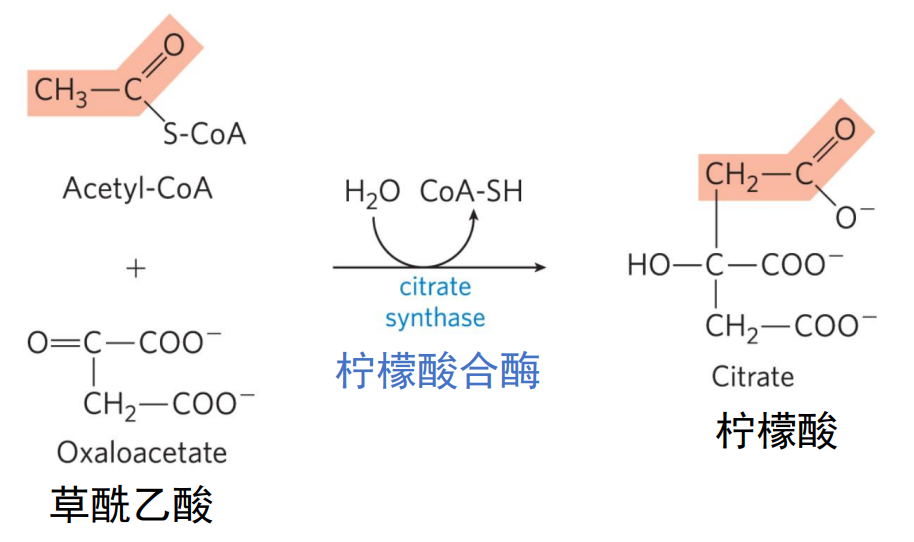
柠檬酸合酶的催化下,乙酰CoA上的甲基碳作为亲核试剂进攻草酰乙酸α-酮基上缺电子的羰基碳,形成柠檬酰CoA中间体。
之后高能碳硫键迅速水解,形成柠檬酸,\(\ce{\Delta G^{'0}=-32.2KJ/mol}\),使得反应不可逆。
柠檬酸合酶通过两个组氨酸与一个天冬氨酸实现催化作用:
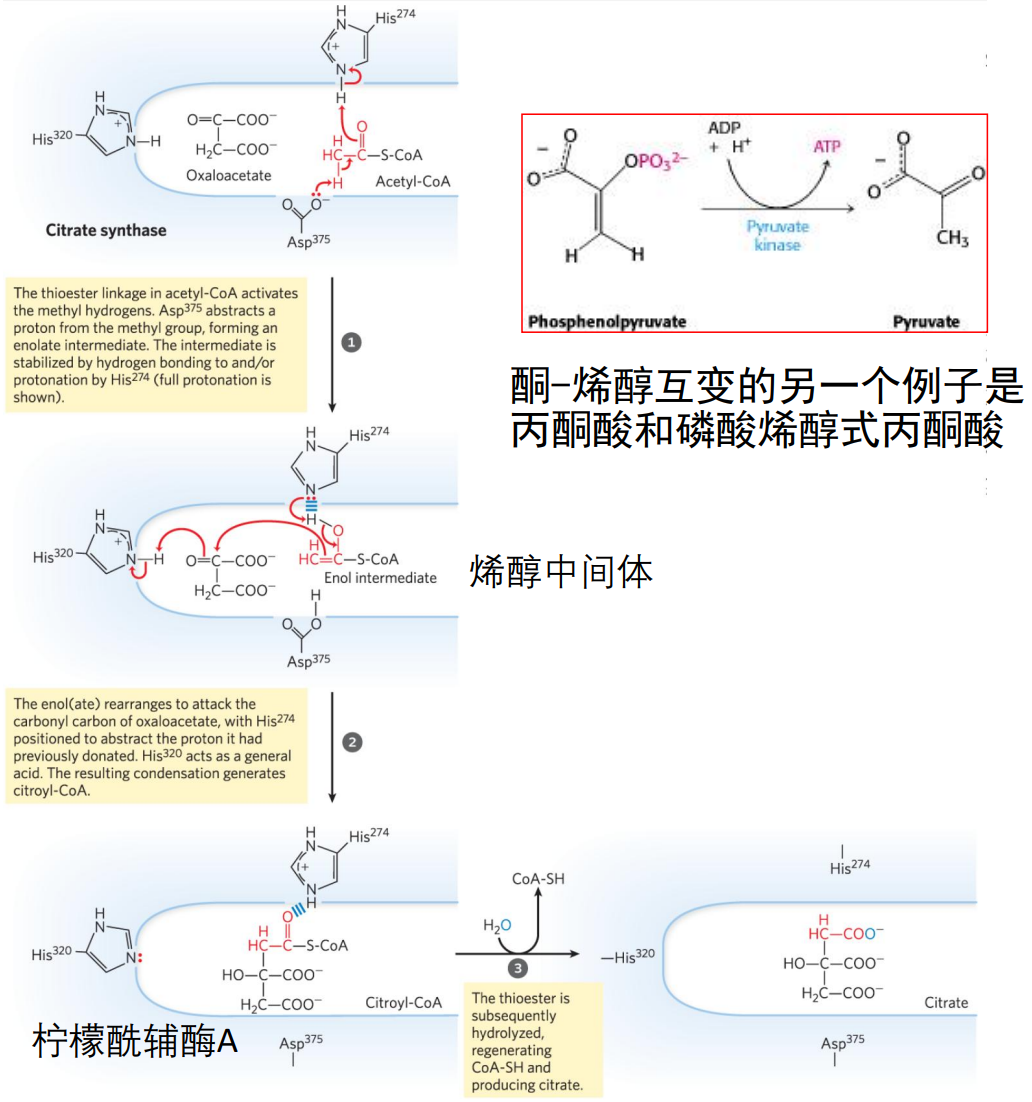
柠檬酸合酶是三羧酸循环的关键调节酶,其催化的反应不可逆,且对草酰乙酸的\(K_m\)极小。
α-酮酸
α-酮酸是一种羧基直接连在α碳上的酮酸,丙酮酸本身是由一个最简单的α-酮酸。
丙酮酸去除甲基部分,为一个草酰基。
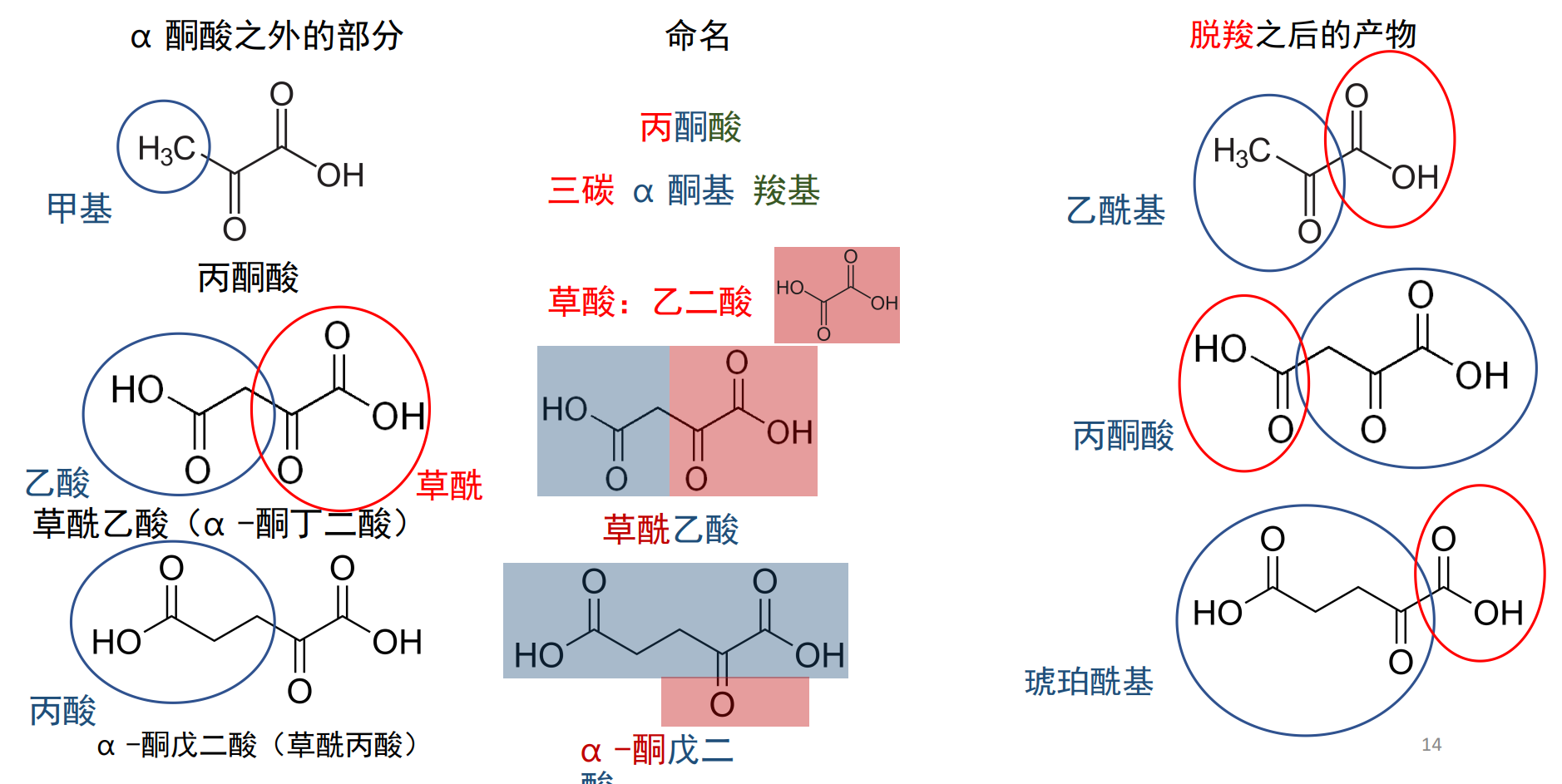
大部分α-酮酸脱羧脱的是草酰基的羧基。(除了草酰乙酸)
第二步 异柠檬酸
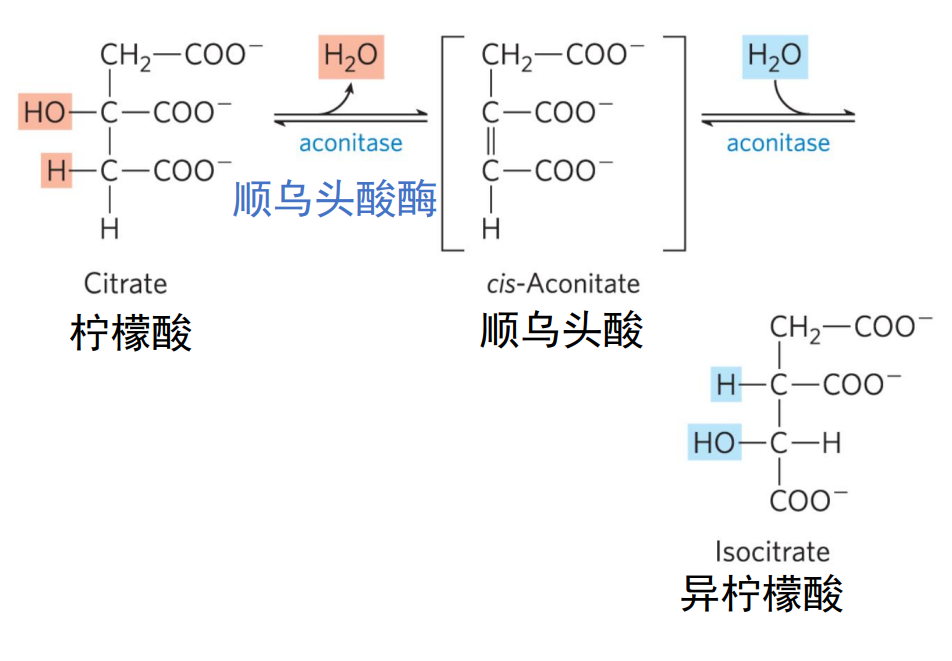
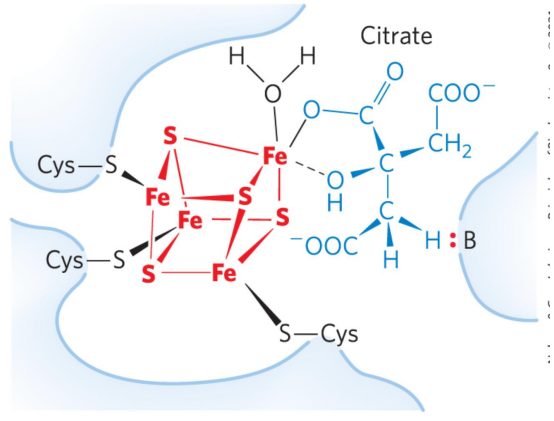
乌头酸酶催化对称的柠檬酸的脱水,再水合,生成不对成的异柠檬酸,本步为可逆反应。
乌头酸酶通过铁硫中心催化反应进行,其是常见的电子载体辅基(复合物\(\rm{I-III_2}\))。
Note
本步反应的脱水具有选择性,双键不能生成在直接来自乙酰CoA的乙酰基上。
第三步 α-酮戊二酸
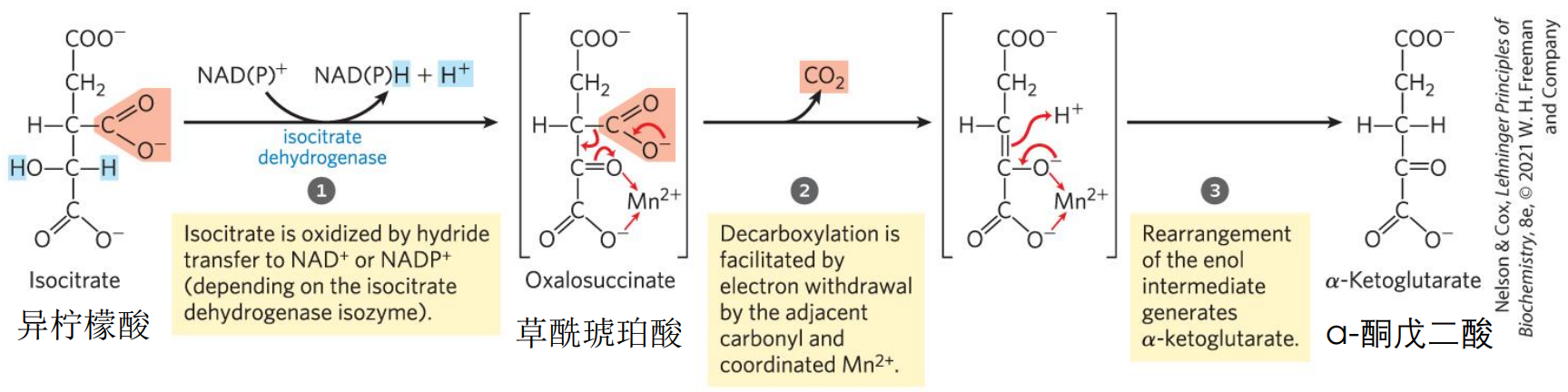
异柠檬酸脱氢酶催化六碳的异柠檬酸脱羧生成五碳的α-酮戊二酸,给出两个电子,\(\ce{NAD+}\)或\(\ce{NADP+}\)作为电子载体接受电子。
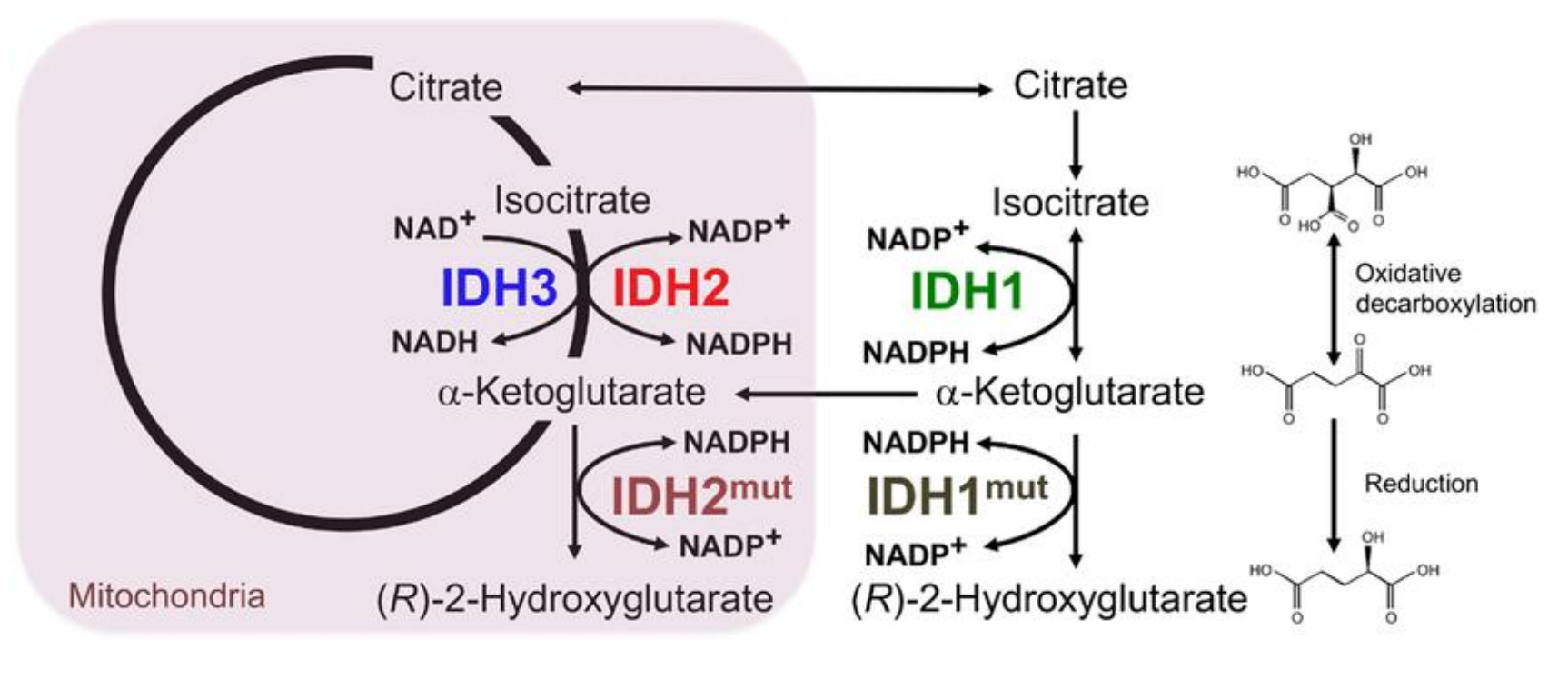
人体内的异柠檬酸脱氢酶有三种同工酶,分别是:
- IDH1,定位于细胞液,以\(\ce{NADP+}\)作为电子受体
- IDH2,定位于线粒体基质,以\(\ce{NADP+}\)作为电子受体
- IDH3,定位于线粒体基质,以\(\ce{NAD+}\)作为电子受体
Note
异柠檬酸脱氢酶突变体会催化生成致癌代谢物。
第四步 琥珀酰CoA
α-酮戊二酸脱氢酶复合物 (α-KGDH or OGDHc) 将α-酮戊二酸氧化脱羧,与辅酶A生成琥珀酰CoA,该步反应是不可逆反应,也是三羧酸循环的重要调节位点。
第五步 琥珀酸
琥珀酰CoA合成酶由两种同工酶,均催化琥珀酰CoA水解生成琥珀酸,但是高能硫酯键能量去向不同,一个走向ATP,一个走向GTP.
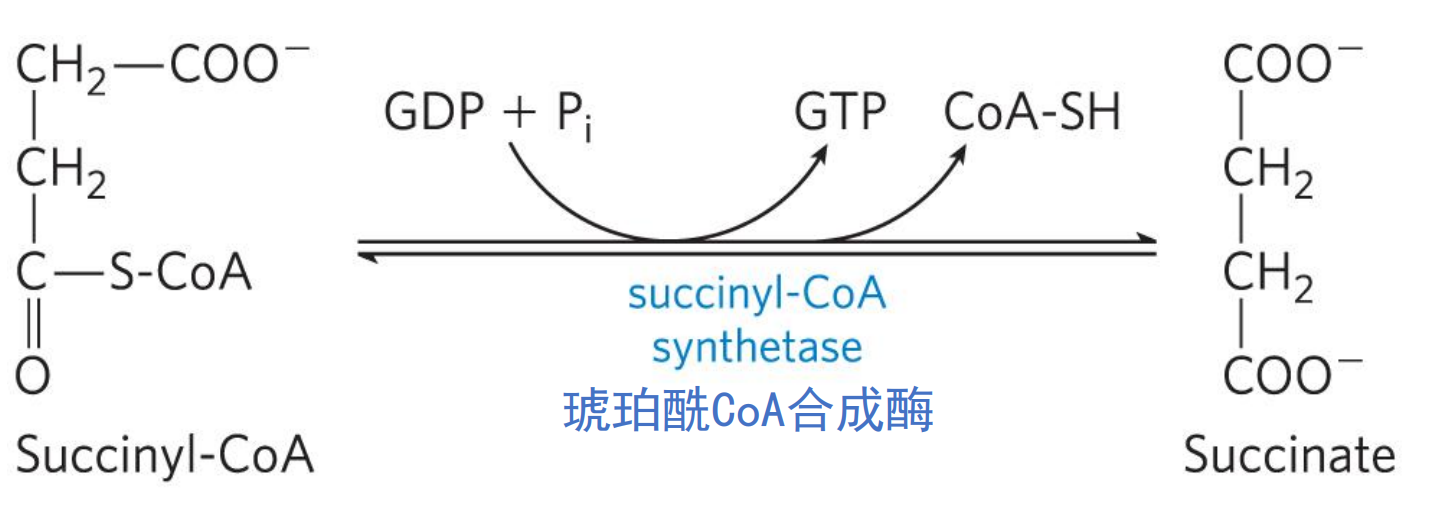
核苷二磷酸激酶 (nucleoside diphosphatekinase) 催化ATP和GTP间的可逆转化。
第六步 延胡索酸
琥珀酸脱氢酶是唯一一个定位于线粒体内膜上的三羧酸循环催化酶,催化琥珀酸脱氢生成延胡索酸:
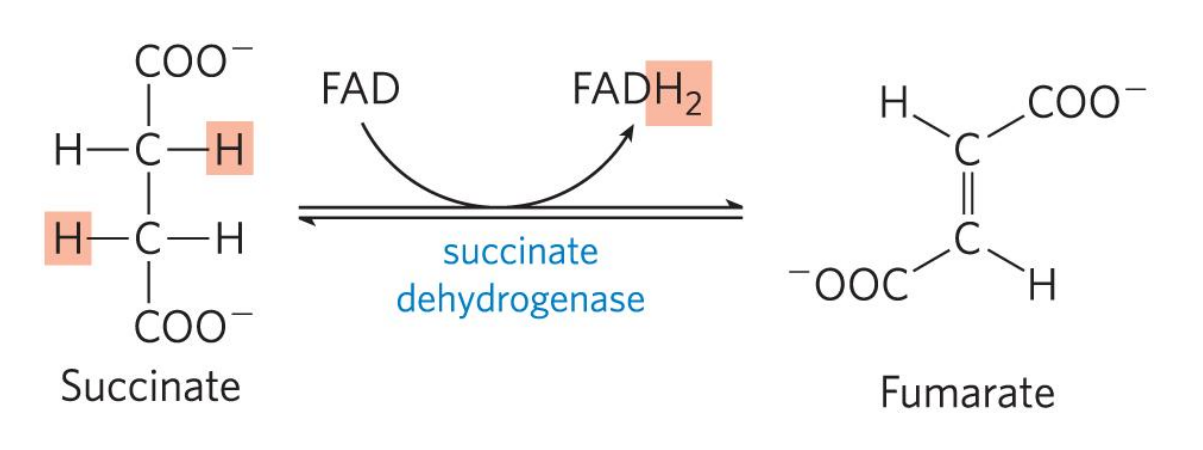
琥珀酸脱氢酶复合体同时也是电子传递链中的复合体II,氧化磷酸化阻断(缺血)时会导致琥珀酸大量积累、
其电子受体为黄素腺嘌呤二核苷酸 (FAD),通过共价键与琥珀酸脱氢酶结合。
辅酶与辅基
辅酶与酶的连接是松散的,辅基与酶的连接是紧密的。
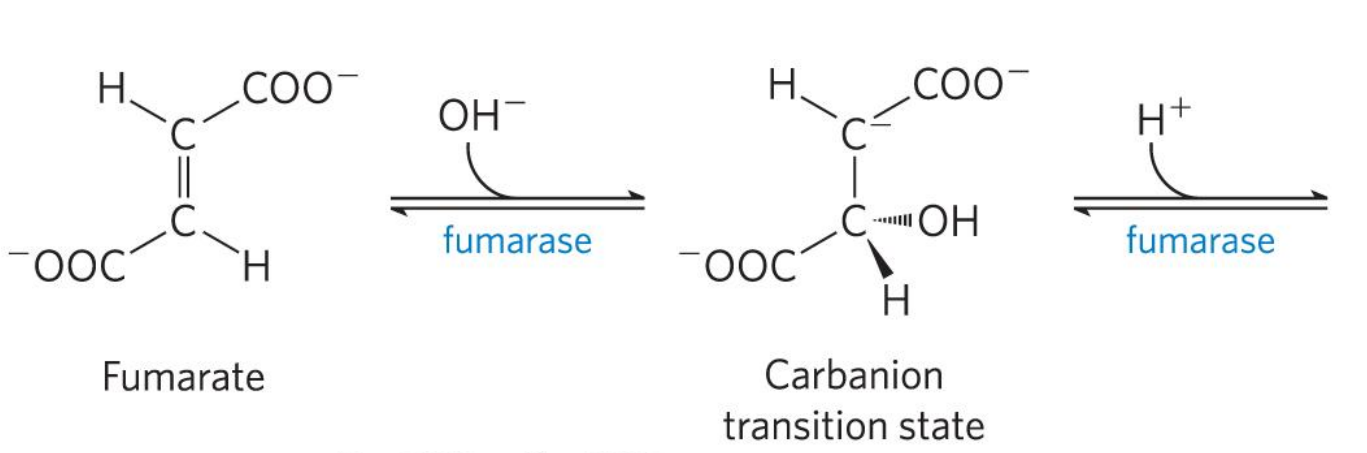
第七步 L-苹果酸
延胡索酸 (反-丁烯二酸)在延胡索酸酶的催化下,以高度的立体专一性,得到左旋的L-苹果酸。该反应是可逆的。
第八步 L-苹果酸氧化生成草酰乙酸
该反应由 L-苹果酸脱氢酶 催化,氧化苹果酸为草酰乙酸,偶联生成NADH。
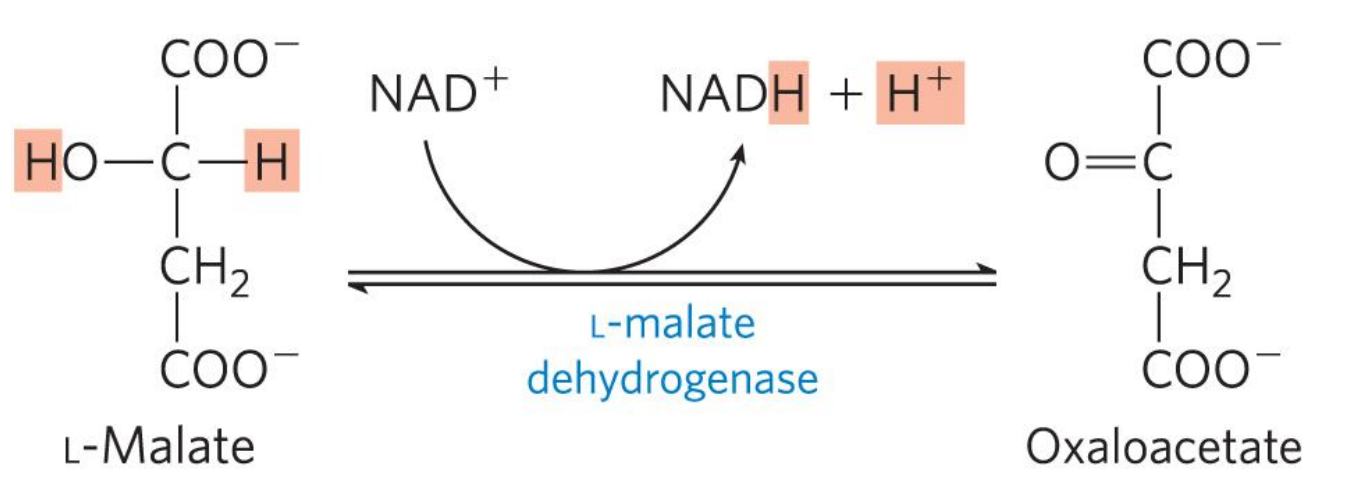
该反应不自发进行,但是线粒体中草酰乙酸浓度极低(柠檬酸合酶\(K_m\)极低,在极低浓度下既可以催化柠檬酸生成)。
三羧酸循环中碳原子命运
一开始柠檬酸是对称分子,但是后续生成异柠檬酸的羟基变位发生在远离来自乙酰CoA的一端。
后续两步异柠檬酸脱羧生成α-酮戊二酸,α-酮戊二酸脱羧生成琥珀酰CoA,两个碳原子都是远离乙酰CoA的一端,均不是来自于乙酰CoA的碳。
但是自琥珀酰CoA生成琥珀酸(丁二酸)开始,变为对称分子,且琥珀酸脱氢酶复合体不存在空间选择性,使得来自乙酰CoA的不对称性湮灭。
三羧酸循环反应总公式
- 2C单位进入,2个CO2离开
- 一分子乙酰CoA,经过循环,产生一分子GTP。
- 从碳能源中收获大量高能电子:3 NADH+1 FADH2。
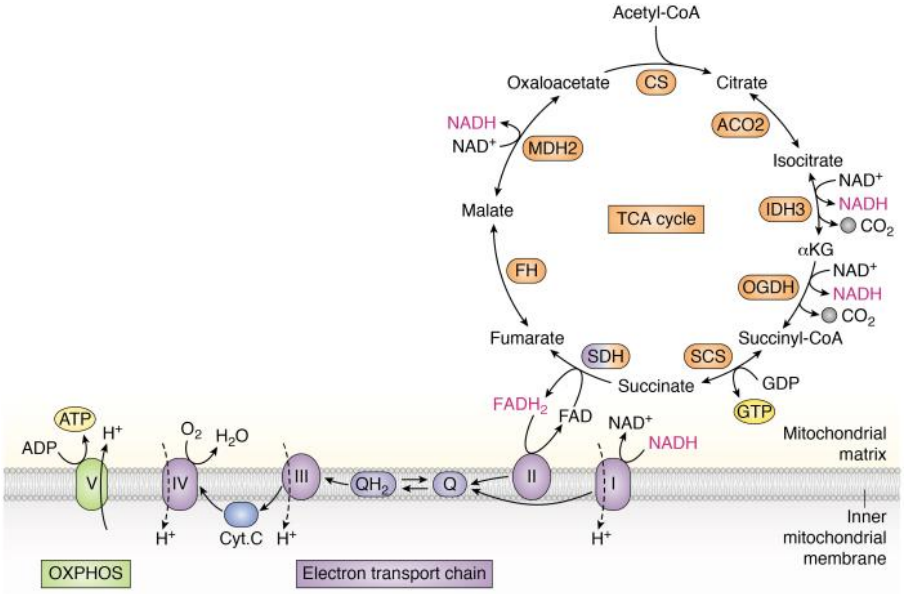
三羧酸循环的生化地位
循环中的代谢前体
下图展示了三羧酸羧酸循环中的重要代谢前提物质:
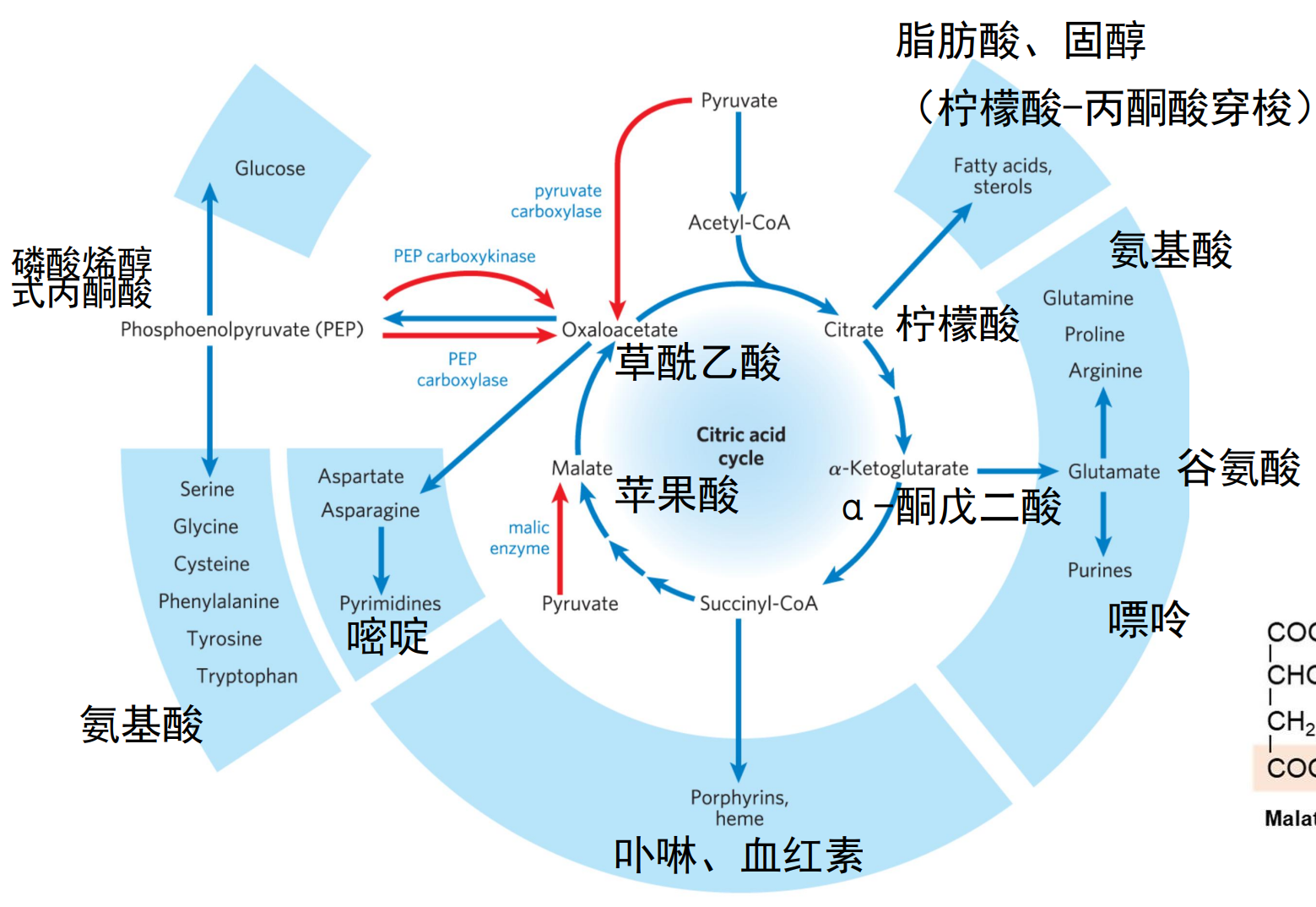
- 柠檬酸参与脂肪酸和固醇的合成
- α-酮戊二酸参与谷氨酸的合成,后者是嘌呤与许多非必需氨基酸的前提物质
- 琥珀酰CoA是卟啉与血红素的前体
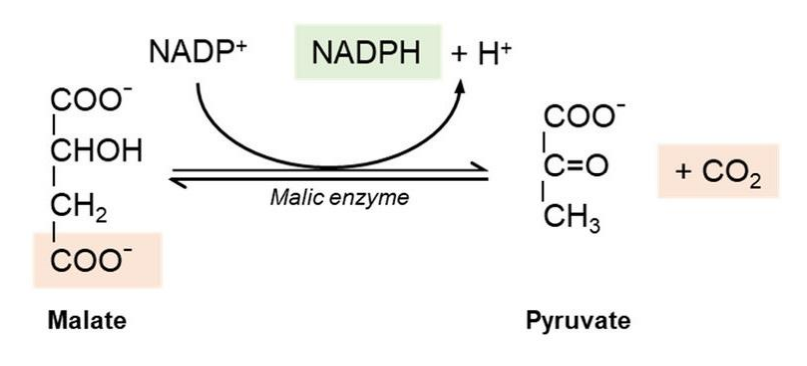
- 苹果酸与丙酮酸之间通过苹果酸酶催化转化
- 草酰乙酸是嘧啶的前体,可以转化为磷酸烯醇式丙酮酸 (PEP),再合成为许多非必需氨基酸与葡萄糖。
因为草酰乙酸的核心地位,三羧酸循环中涉及的所有分子都是生糖的。
添补反应
由于草酰乙酸在线粒体内浓度很低,三羧酸循环本身的通量很小,反应路径上的大部分分子的存货都不算多。
为了保证三羧酸循环高效进行,细胞必须为其准备一些“后备能源”,保证某一个环节“缺货”时可以被及时补充。这种对参与其他代谢途径而消耗的TCA循环中间产物的及时补充,保持TCA循环顺利进行的反应,被称为添补反应。
最重要的添补反应——由丙酮酸羧化酶催化的从丙酮酸生成草酰乙酸的反应。
草酰乙酸添补反应
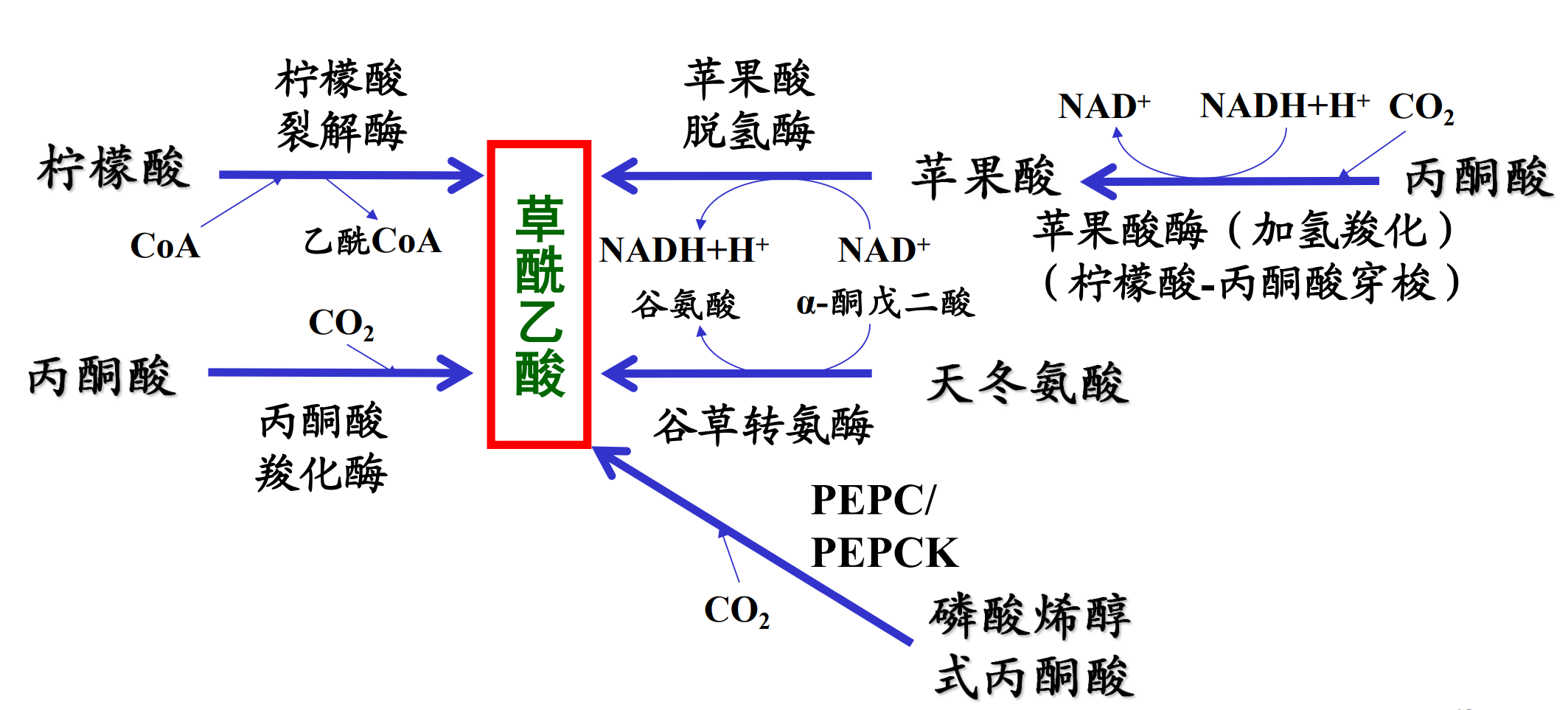
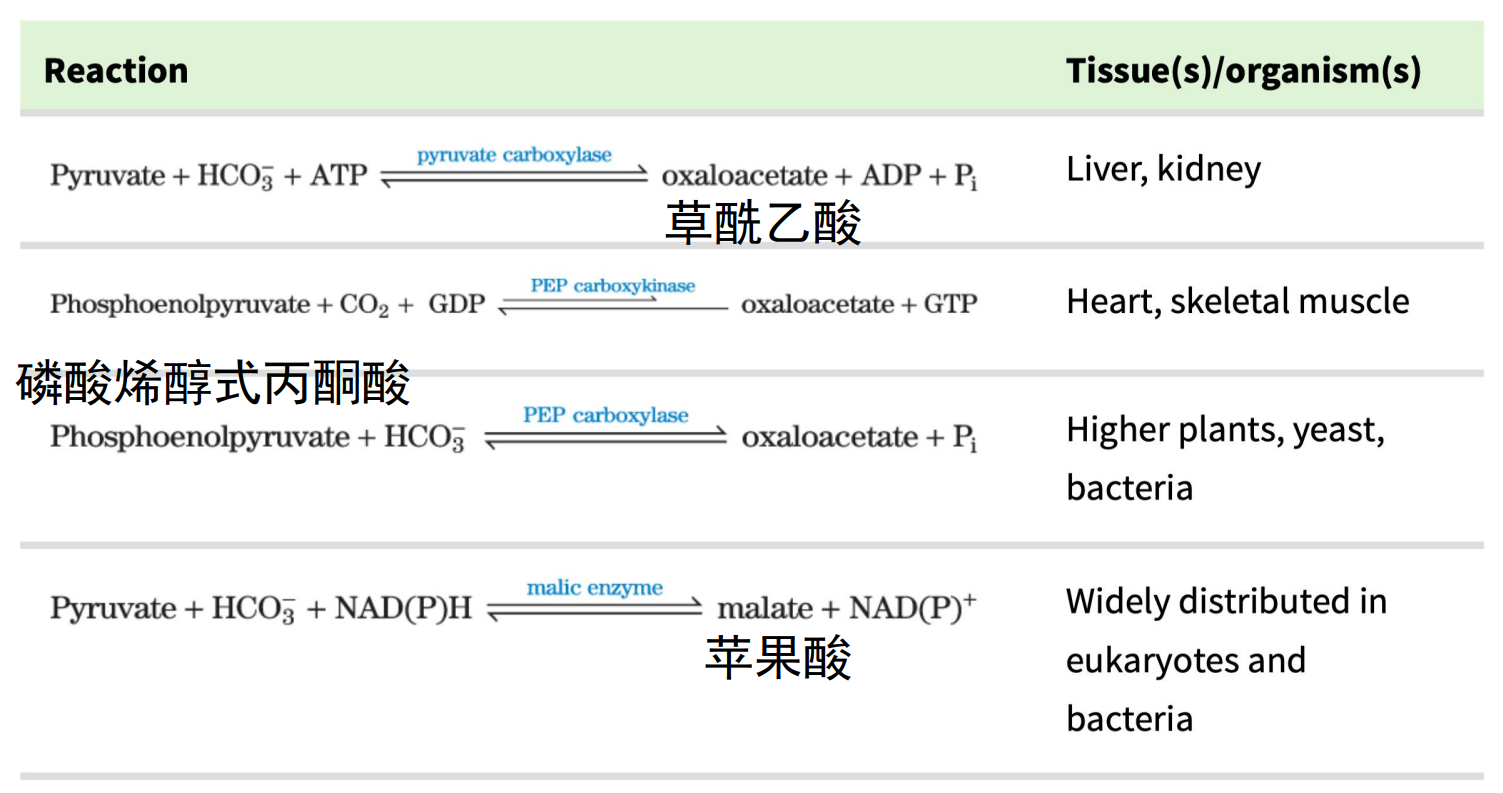
如果有大量的乙酰-CoA却没有足够的草酰乙酸(在以脂肪酸为主要能源时)会大量生成酮体。
三羧酸循环生理意义
- 糖、脂肪、蛋白质的最终代谢通路
- 糖、脂肪酸、氨基酸代谢联系的枢纽
- 糖、脂肪酸、氨基酸及其它物质合成的前体物质的重要来源
- 与氧化磷酸化共同为需氧细胞提供绝大多数的能量
三羧酸循环调控
三羧酸循环有多个关键调节位点:
- 底物、产物对循环中的三个催化不可逆反应的关键酶产生影响:柠檬酸合酶,异柠檬酸脱氢酶(生成CO2),α-酮戊二酸脱氢酶(生成CO2)。
- 上、下游的糖酵解、氧化磷酸化都对其有重要调节作用