信号转导
细胞通讯与细胞外信号
细胞通讯 (Cell communication),指的是一个细胞发出的信息、通过介质(又称配体)传递到另一个靶细胞并与其相应的受体相互作用,然后通过细胞信号转导产生靶细胞内一系列生理生化变化,最终表现为靶细胞整体的生物学效应的过程。
信号传导 (signal transduction),指的是胞外信号分子与细胞膜上或胞内的受体特异性结合,将信号转换后传给相应的胞内系统,使细胞对外界信号做出适当的反应的过程。
细胞通讯的方式
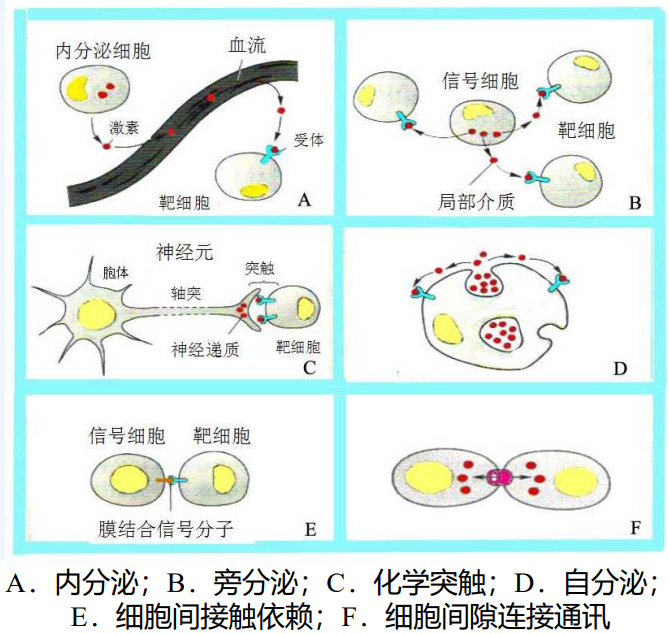
细胞间通讯方式可粗略分为三种:分泌化学信号进行细胞间通讯、细胞间接触依赖性通讯 (contact-dependent signaling) 与动物细胞间形成的间隙连接 (gap junction)。
分泌化学信号
细胞在分泌化学信号上,根据不同的通讯需求,有不同的分泌方式:
- 内分泌 (endocrine signaling):信号分子通过血液循环运输到身体各处,作用远距离的靶细胞
- 旁分泌 (paracrine signaling):局部化学介质经过细胞外液中局部扩散 ,作用于邻近靶细胞。比如通过化学突触分泌神经递质传递神经信号。
- 自分泌 (autocrine signaling):细胞分泌的信号分子作用于自身细胞。
接触依赖性通讯
这种方式需要细胞间“贴贴”,通过直接接触,使得细胞跨膜信号分子(配体)与相邻靶细胞表面受体相互作用。
其包括细胞-细胞黏着和细胞-基质黏着两种机制,特别地,细胞间接触依赖性通讯参与胚胎发育过程中对组织内相邻细胞的分化命运的决定。
Note
接触依赖性通讯的过程中无需信号分子的释放。
间隙连接通讯
如果说接触依赖性通讯是“贴贴”,那间隙连接通讯就是“亲嘴”了!它是一种特化的细胞-细胞连接,通过水溶性通道直接连接相连细胞的胞质,通过交换小分子来实现代谢偶联或电偶联,是细胞直接交流信息的方式。
细胞通讯的过程
无论如何,细胞通讯的一个基本模型大致如下:
- 产生信号的细胞合成并释放信号分子
- 运送信号分子至靶细胞
- 信号分子与靶细胞受体特异性结合并导致受体激活
- 活化受体启动胞内一种或多种信号转导途径
- 引发细胞功能、代谢或发育的改变
- 信号的解除并导致细胞反应终止
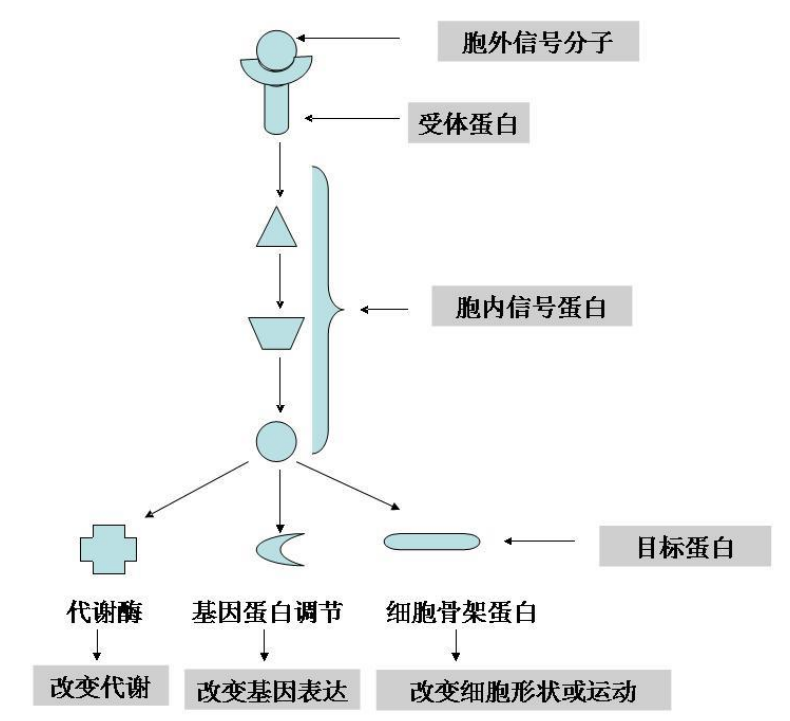
万变不离其宗,其中最核心的是后四步,它们涉及到三个问题:胞外信号是如何传入胞内的、细胞通讯是如何影响靶细胞生理状态的与这个信号是如何终止的。
细胞外信号分子
信号分子 (signal molecule) 是细胞的信息载体,种类繁多,包括化学信号以及物理信号等。其中最重要的是由细胞分泌的、能够调节机体功能的一大类生物活性物质,它们是细胞间通讯的信号,被称为第一信使。
根据溶解性,可以分为亲脂性信号分子——胞内受体,亲水性信号分子——胞外受体。
根据作用方式,可以分为激素——远距离作用,神经递质——作用突出后膜,局部化学介质——就地解决。
根据作用后效应可以分为激动剂——与受体结合刺激效应的产生,拮抗剂——阻碍激动剂作用。
受体
- 受体 (receptor) :存在于胞膜或胞内的大分子多(为蛋白质),能特异性识别并结合胞外信号分子,进而激活胞内一系列生物化学反应,使细胞对外界刺激产生相应的效应。
- 配体 (ligand) :是与受体结合的生物活性物质的统称,包括激素、神经递质、生长因子、某些药物和毒物等。
受体根据其所固定在细胞中的位置分为两类:
- 膜受体:镶嵌在胞膜上的糖蛋白、糖脂。
- 结构:细胞外域——与配体相互作用,跨膜域——将受体固定在细胞膜上,胞内域——起传递信号作用。
- 配体:肽类激素、生长因子与递质等亲水分子。
- 胞内受体:
- 核受体:DNA结合蛋白,作为转录因子与DNA顺式作用元件结合,调节基因的表达。其配体是来自胞外的亲脂性小分子如甾类激素。
- 胞浆受体:一氧化氮受体与一氧化碳受体。
膜受体
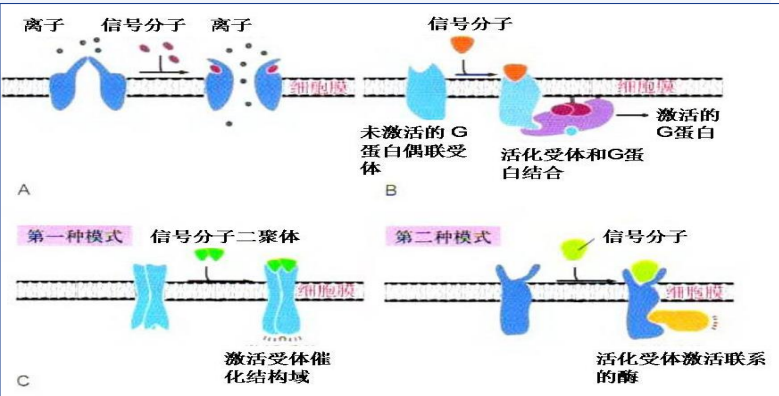
膜受体主要包含离子通道偶联受体、G蛋白偶联受体与酶联受体。
 、
、
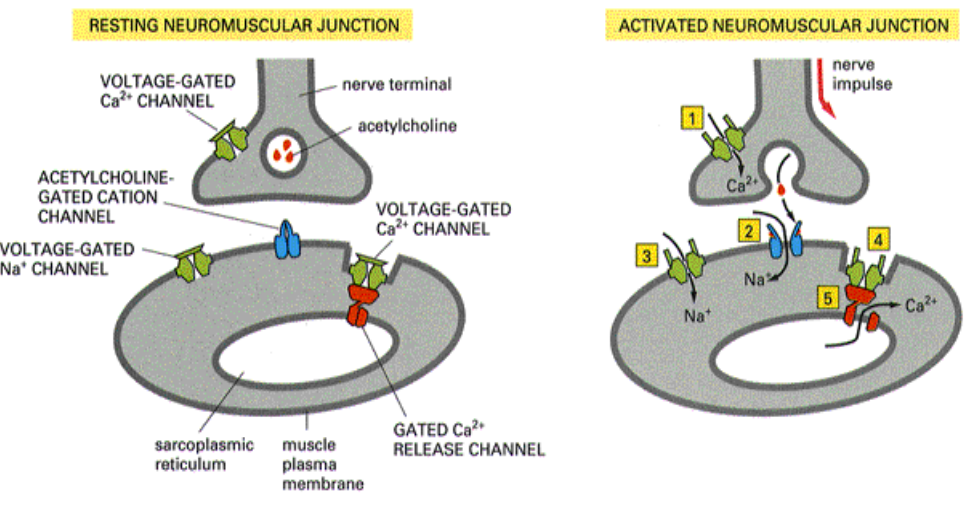
离子通道偶联受体
离子通道偶联受体 (ion channel receptor) 既可与细胞外信号分子结合,同时又是离子通道,因此具有受体与离子通道偶联的特点。介导的信号转导反应是一种快速的反应,为神经系统和其他电激发细胞所特有由多个亚基组成的多聚体,每个亚基具有2、4、5或6个跨膜域。
Note
简单来说,离子通道偶联受体身兼两职——受体与离子通道,且两者偶联,相当于包装了两个功能。抽象!
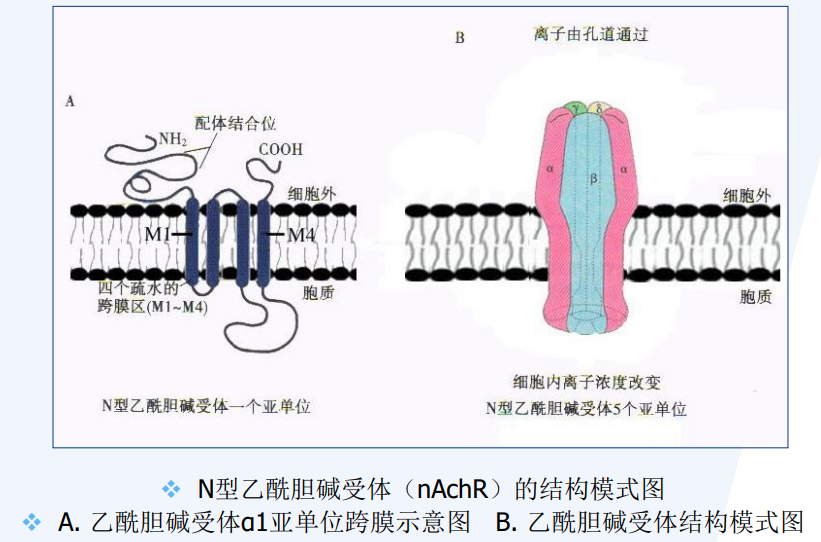
离子通道偶联受体总体上可分为两个超家族:
- I型受体超家族:每一亚基各含2、4或5个跨膜区域,通过其胞外区域与配体结合。
- Ⅱ及Ⅲ型受体超家族:每一亚基均有6个跨膜区域,受体与配体的结合部位在细胞膜,而不是在胞外。
突触后膜上常见离子通道偶联受体。
G蛋白偶联受体
G蛋白偶联受体 (G protein linked receptor) 是膜受体中最大的家族,真正的王朝。虽然其介导的信号转导过程较慢,但敏感、灵活,而且类型多样。
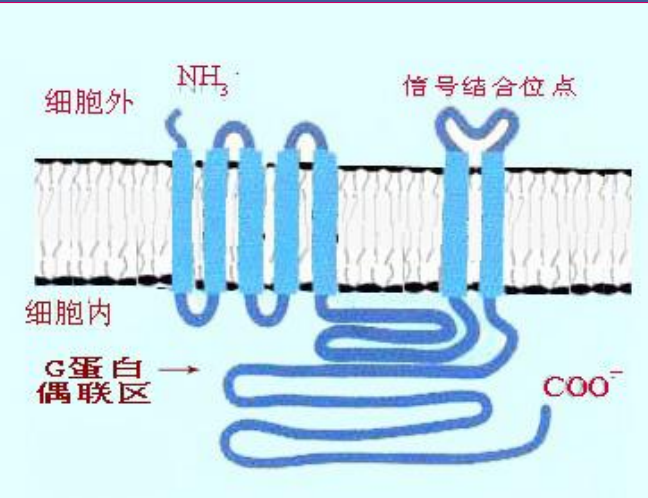
G蛋白偶联受体为为一条多肽链构成的糖蛋白,分为:
- 胞外区:N末端,带有多个糖基化位点。
- 包膜结构区:标志性的7个跨膜α螺旋结构,氨基酸组成高度保守,各跨膜螺旋结构之间有6个环状结构(胞外、胞内各三个)。
- 胞内区:C末端,该位点的丝氨酸、苏氨酸为磷酸化部位。
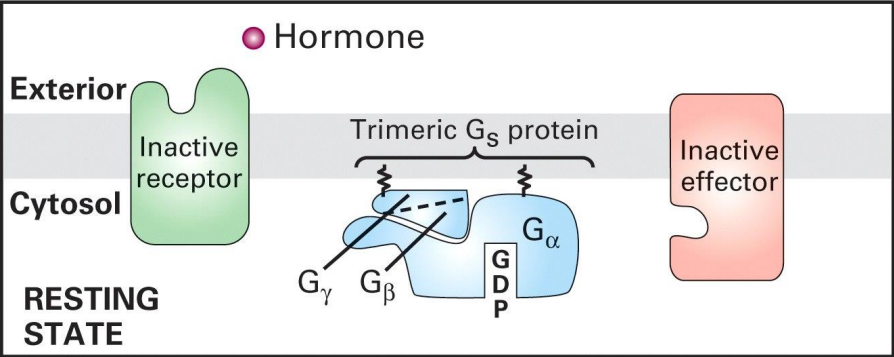
G蛋白
G蛋白,指的是在信号传导中,与受体偶联,且能与鸟苷酸 (GMP) 结合的三体GTP结合蛋白,可溶性外周蛋白,具有α、β和γ三种蛋白亚基。
G蛋白不是G蛋白偶联受体!!!
其通过自身构象变化激活效应蛋白 (effector protein),实现胞外信号传递至胞内。
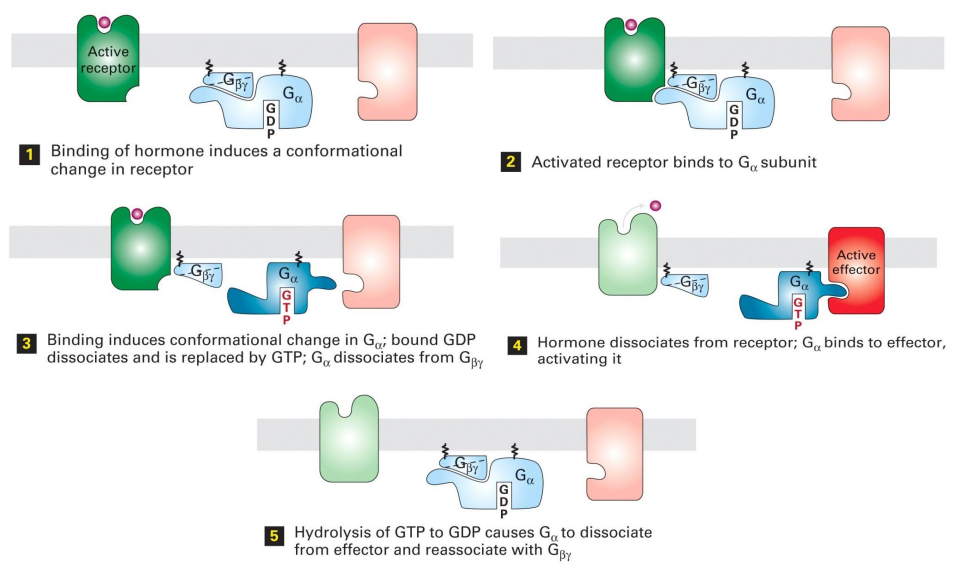
G蛋白的活化过程需要G蛋白偶联受体的参与:当配体与G蛋白偶联受体结合时候,受体分子构象改变,接触并改变G蛋白α亚基构象,使其摆脱GDP,结合新的GTP,同时α亚基的构象改变使得其“抛开”β、γ二聚体,前者激活下游效应蛋白,后者在膜上自由扩散。
而当配体与受体解离,G蛋白α亚基水解GTP为GDP构象改变,重新与β、γ二聚体结合,回到静息状态。
G蛋白的下游效应蛋白包括腺苷酸环化酶(AC)、磷脂酶C(PLC)等。(由于G蛋白固定在膜上,通常是膜上的酶或离子通道)
根据G蛋白特点与作用,可以分为:
- 激动型G蛋白(Gs家族):激活腺苷酸环化酶
- 抑制型G蛋白(Gi家族):抑制腺苷酸环化酶
- 磷脂酶C型G蛋白(Gq家族):激活磷脂酶C
- ...
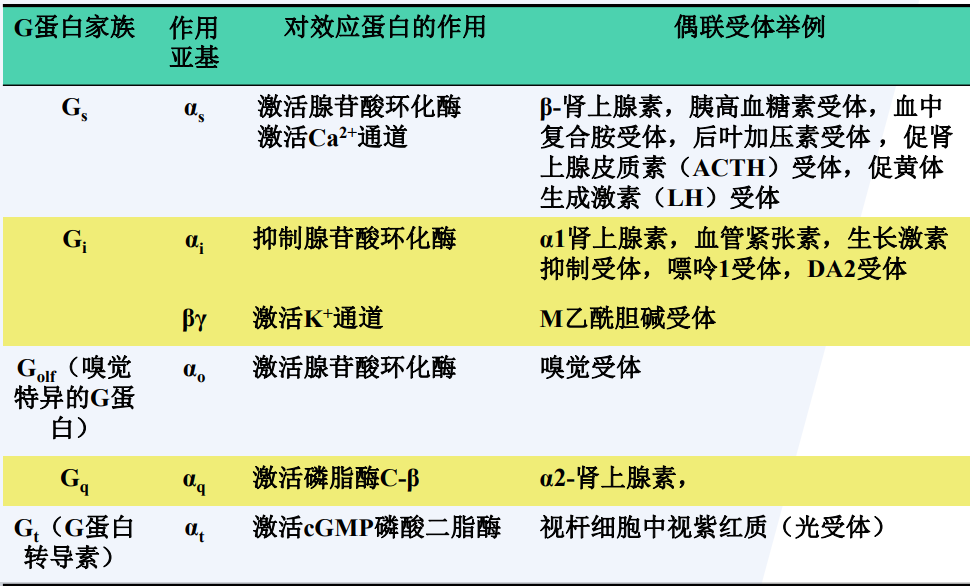
酶联受体
酶联受体是第二大类的细胞表面受体,也是一类跨膜蛋白,配体结合结构域位于胞外侧,其要么本身就具有酶活性,要么就与某种酶深度绑定。
酶联受体的每个亚基通常只有一个跨膜片段。
酶联受体分为五类:
- 受体酪氨酸激酶
- 酪氨酸激酶关联受体
- 受体类似的酪氨酸磷酸酶
- 受体丝氨酸/苏氨酸激酶
- 受体鸟苷酸环化酶
受体酪氨酸激酶 RTK | 酪氨酸蛋白激酶型受体 RTPK
RTK 是一条多肽链构成的跨膜糖蛋白,具有酪氨酸激酶活性:
- 胞外区:N端,是配体结合部位。
- 胞质区:C端,含酪氨酸激酶功能区,该区在氨基酸组成上高度保守,包括结合ATP与结合底物两个区域
- 跨膜区:由一个高度疏水的α螺旋构成,由22~26个氨基酸组成。
七种亚家族的受体酪氨酸激酶:
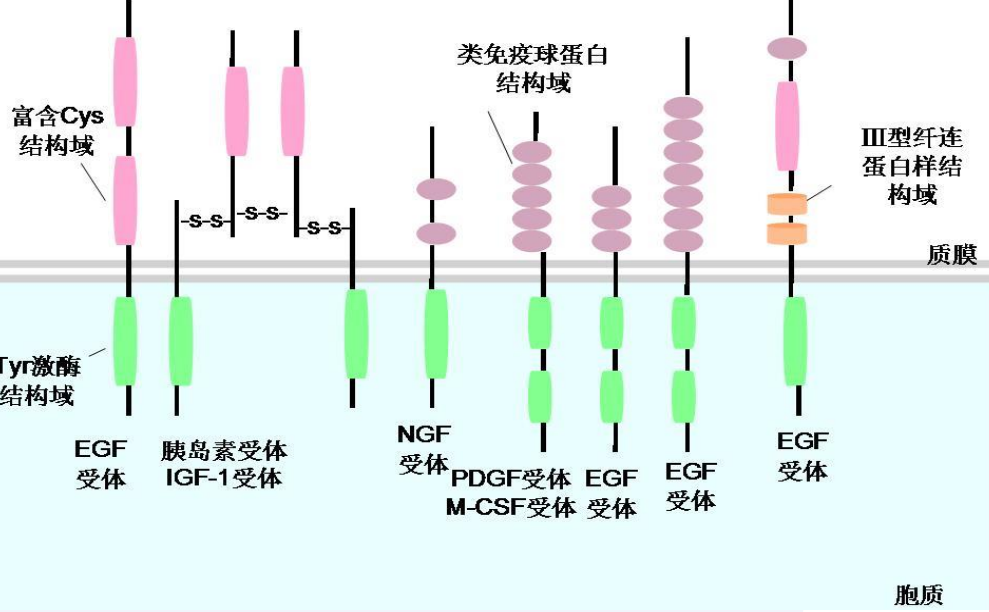
下图展示了配体结合所诱导的受体酪氨酸激酶的二聚化与自磷酸化:
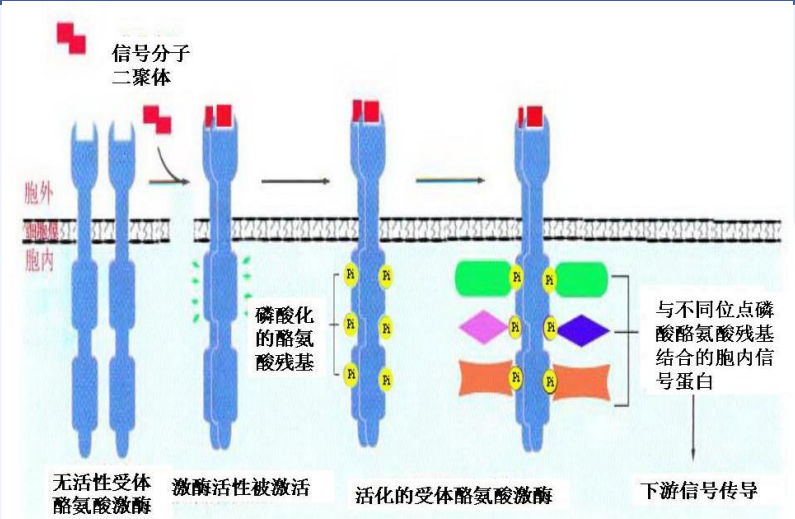
RTK 的作用过程大致为:配体与 RTK 结合后,受体胞外结构域构象改变,激发二聚化,之后 RTK 活性增强,发生C端酪氨酸残基磷酸化,也就是受体自磷酸化,诱发形成 SH2 结合位点,招募具有 SH2 结构域的胞内信号蛋白,进一步向下传导信号。
RTK 的主要配体为一些生长因子和分化因子,如表皮生长因子EGF)、血小板源生长因子(PDGF)和胰岛素(insulin)等,在参与细胞生长和分化的调控,引起细胞产生效应的过程较缓慢。
酪氨酸激酶关联受体 (tyrosine kinase-linked receptor)
它就像是一个盗版 RTK,其具有酪氨酸蛋白激酶的结合位点,但本身不具有酪氨酸蛋白激酶活性,需要非受体酪氨酸蛋白激酶的参与。
其配体为造血系统中调节细胞增殖与分化的局部介质(细胞因子)、某些激素如生长激素和催乳素以及T淋巴细胞和B淋巴细胞抗原等。
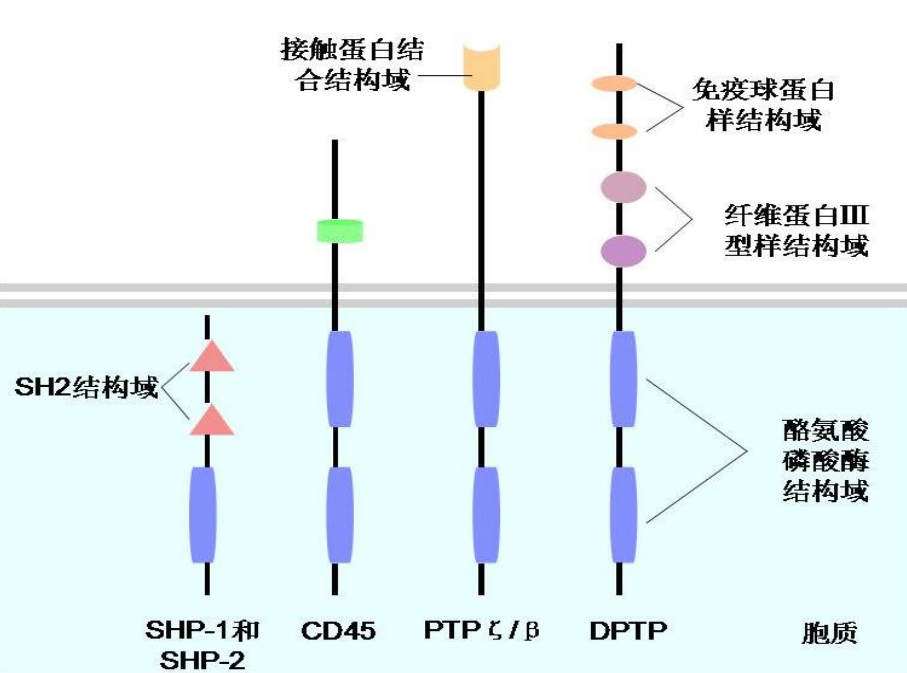
受体类似的酪氨酸磷酸酶 (receptor tyrosine phosphatase)
它则与 RTK 反着来,作为一次跨膜蛋白受体,胞内区具有蛋白酪氨酸磷酸酶活性,配体结合时候激发活性,使得胞内信号蛋白 的磷酸酪氨酸残基脱磷酸化。
其主要参与维持静息细胞内酪氨酸残基低磷酸化,在细胞信号系统中发挥特殊的调节作用,在细胞周期调控中发挥重要作用。
下图展示了几种受体类似的酪氨酸磷酸酶:
受体丝氨酸 / 苏氨酸激酶 (receptor serine / threonine kinase)
它是一种二聚体蛋白,由I型受体亚单位和II型受体亚单位组成,其胞质侧具有丝氨酸和苏氨酸激酶活性,配体以蛋白二聚体的形式与其结合,使激酶活化。
受体丝氨酸/苏氨酸激酶是一类生长因子受体,配体包括激活素、抑制素、骨形态发生蛋白和转化生长因子等,参与基因表达的调控。
受体鸟苷酸环化酶 (receptor guanylate cyclase)
受体鸟苷酸环化酶是动物细胞中存在的一次跨膜蛋白受体,胞外段是配体结合域,胞内段为鸟苷酸环化酶催化结构域,具有鸟苷酸环化酶活性,分解GTP为cGMP,从而激活下游靶蛋白(比如一些依赖cGMP的蛋白激酶)。
胞内受体
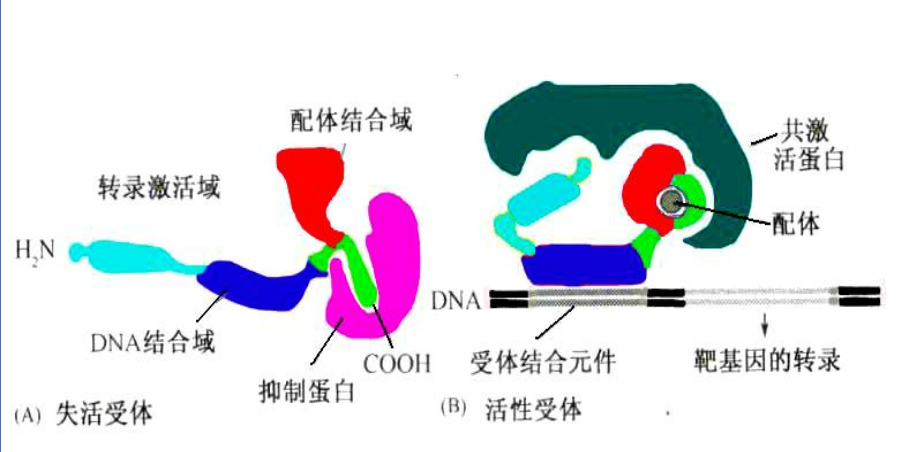
核受体
这类受体与配体结合激活后都将入核,与DNA结合,激活基因表达,故称为核受体。主要分布在胞浆或细胞核内,通常为400~1000个氨基酸组成的单体蛋白。
其配体一般是脂溶性小分子甾体类激素,以类固醇激素类较为常见,比如甾类性激素、皮质醇、维生素D、类视黄素等。此外还包括甲状腺激类激素等。
核受体一般结构如下:
- N端转录激活结构,有一定转录激活作用,序列高度可变。
- DNA结合结构域:序列高度保守,具有两个锌指结构,与DNA结合。
- 铰链区域:链接作用
- C端配体结合域:与配体结合,引起受体活化。
核受体与配体结合活化后,引起分子构象改变,进而与DNA序列上被称为激素调节元件 (HRE) 结合,从而稳定/干扰转录因子与DNA序列的结合,选 择性地促进或抑制基因转录。
核受体引起细胞产生效应的速度较慢,一般需要数小时至数天,其包含主要的两个阶段:
- 快速的初级反应阶段:直接激活少数特殊基因转录。
- 延迟的次级反应阶段:初级反应的基因产物再激活其他基因转录,对初级反应起到了放大作用。
胞浆受体
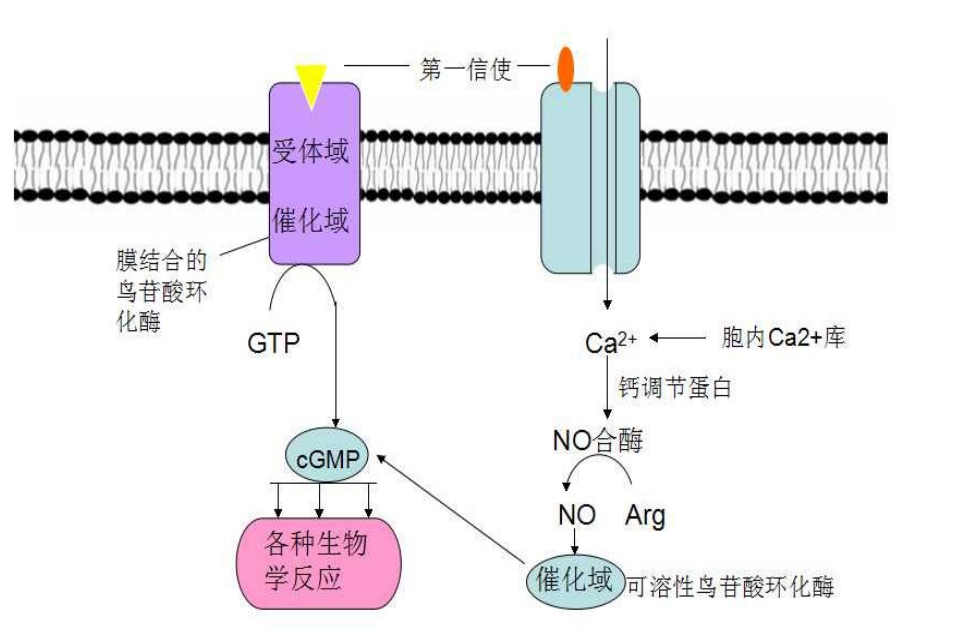
胞浆受体的典型例子就是细胞内具有鸟苷酸环化酶 (G-cyclase, Gc) 活性的 NO 受体和 CO 受体。
以 NO 受体举例,内源性 NO 直接扩散穿膜来到邻近细胞,与胞内受体鸟苷酸环化酶中心的\(\ce{Fe^{2+}}\)结合,改变后者构象,使得酶活性大大增强与cGMP水平升高,引发下游一系列事件的发生。
受体作用特点
- 受体能选择性地与特定配体结合
- 受体与配体的亲和力强
- 受体-配体结合后显示可饱和性
- 受体-配体的结合具有可逆性
- 受体与配体的结合可通过磷酸化和去磷酸化进行调节
细胞内信使
第二信使 (second messenger),是细胞信号转导过程中的接力选手,大多数为亲水的第一信使服务,因为后者只能在胞外通过膜上受体将信号传递至胞内,而从胞内继续向下传递需要第二信使的接力,在此过程中它们还能一步步放大生物学效应,实现“举一旗,众人皆呼”的号召力。
第二信使的定义为指受体被激活后在细胞内产生的、能介导信号转导的活性物质。主要包括:cAMP、cGMP、二酯酰甘油(DAG)、三磷酸肌醇(IP3)、钙离子、3,4-二磷酸酯酰肌醇[PI(3,4) P2]和3,4,5-三磷酸磷脂酰肌醇(PIP3)等。
接下来让我们来认识一下这里面的明星选手!
cAMP信使体系
环磷酸腺苷 (cyclic AMP, cAMP),是最重要的胞内信使。
由腺苷酸环化酶 (adenylate cyclase, AC) 催化 ATP 脱去 PPi 后自环化生成 cAMP。cAMP 可以在磷酸二酯酶 的催化下水解为 5'-AMP。两步反应都需要\(\ce{Mg^{2+}}\)的参与。
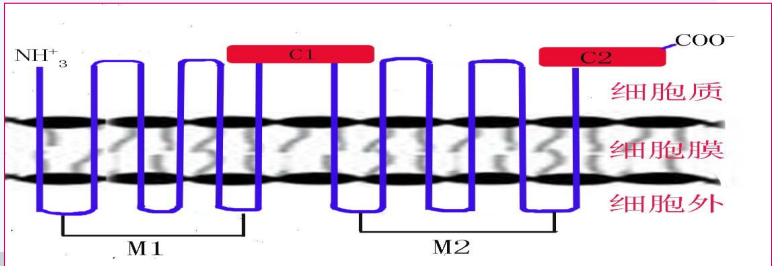
腺苷酸环化酶
AC 是一种膜蛋白,具有两个大疏水区域 (M1, M2),每个均跨膜6次。具有两个高度保守的胞质区域 (C1, C2),是 ATP 结合位点与酶活性部位。
其将 ATP 催化为 cAMP。
cAMP作用机制
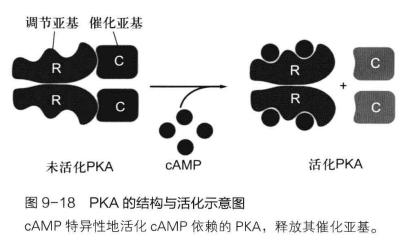
cAMP 将转导信号的接力棒接过来后,需要在细胞内找一个靶向分子来“传达圣旨”,在细胞内,它的最好拍档就是 cAMP依赖性蛋白激酶A (cAMP-dependent protein kinase A, PKA),它是一个能被 cAMP 活化的蛋白激酶,由催化亚基(C亚基)与调节亚基(R亚基)组成 C2R2 四聚体。
每个R亚基上有两个 cAMP 结合位点,这种结合是协同方式的结合,意味着第一个 cAMP 的结合会降低第二个 cAMP 结合的难度(解离常数降低),因此极小的 cAMP 浓度就能触发 PKA 的活性。
当 PKA 的两个R亚基集齐4个 cAMP 时,PKA 活化,它将C亚基以单体形式抛出,而C亚基的蛋白激酶活性被激发,可以对特定序列中的丝氨酸/苏氨酸进行磷酸化修饰,调节特定蛋白的活性,进而将胞外信号转化为胞内的生物学效应。
PKA对底物的特异性要求较低,它基本只要一段符合的氨基酸序列就可以把磷酸基团插在上面,因此催化的底物相当广泛,导致 cAMP 的生物学效应多样。
一个经典的 cAMP-PKA 信号通路大抵如下:
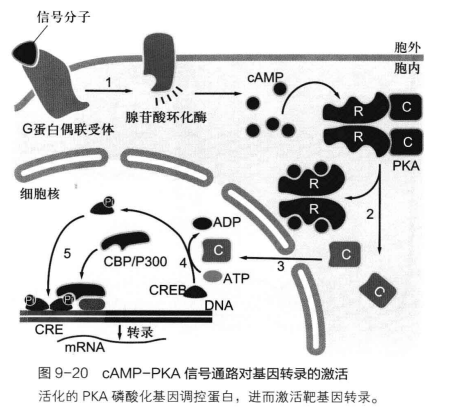
cAMP 除了通过激活 PKA,使得下游蛋白因磷酸化而激活/抑制引发生物学效应,还可以通过调节离子通道通透性来实现信号的向下转导。
cGMP信使体系
环磷酸鸟苷 (cyclic GMP,cGMP),是一种广泛存在于动物细胞中的胞内信使。
由鸟苷酸环化酶 (guanylate cyclase, GC) 催化 GTP 脱去 PPi 后自环化生成 cGMP。cGMP 可以在磷酸二酯酶 的催化下水解为 5'-GMP。两步反应都需要\(\ce{Mg^{2+}}\)的参与,其中后一步反应\(\ce{Ca^{2+}}\)具有类似作用。
下表展示了动物体内不同 GC 的分布:
在同一种细胞中,随着细胞生长过程的变化,两种GC的比例可发生改变。
cGMP作用机制
与 cAMP 类似,cGMP 的“黄金拍档”是 cGMP依赖蛋白激酶G (cGMP depedent protein kinase G,PKG),它是一个二聚体,由催化亚基和调节亚基组成。其主要的催化底物是组蛋白、磷酸化酶激酶、糖原合成酶及丙酮酸激酶等。除此之外,PKG 还可通过磷酸转移酶作用自身磷酸化,通过抑制方式来调节活性。
除了靶向 PKG,cGMP 可以直接作用于阳离子通道,调节其通透性。
IP3/DAG 信使体系
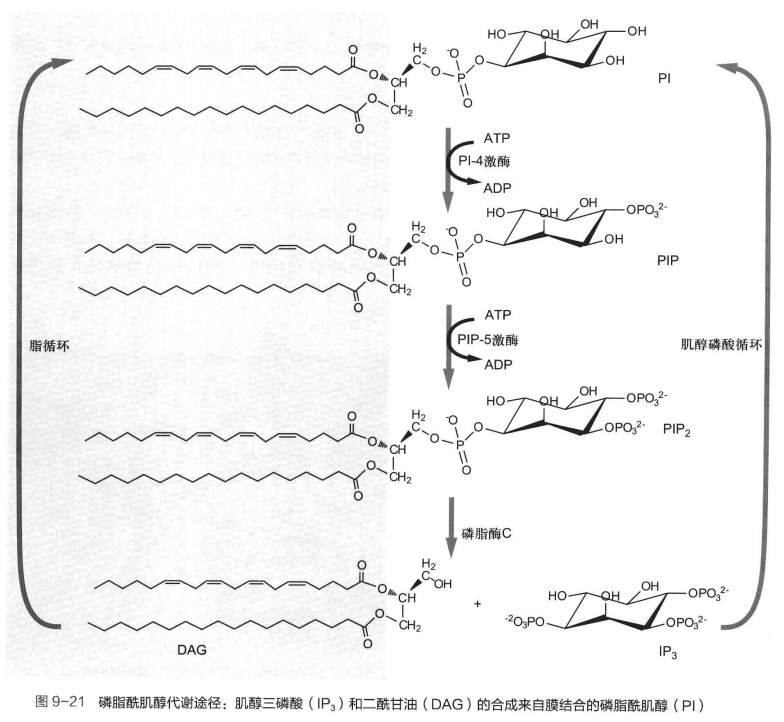
二脂酰甘油 (DAG) 与 三磷酸肌醇 (IP3) 由活化的G蛋白通过激活磷脂酶C (PLC),催化胞膜脂质内层磷脂酰肌醇4,5-二磷酸 (PIP2)水解生成。前者为脂溶性,后者为水溶性。
Note
于磷脂酰肌醇 (PI) 经过两次肌醇环羟基磷酸化生成PIP2。
IP3/DAG 作用机制
IP3 生成后自膜上扩散至胞质中,与内质网膜上IP3敏感\(\ce{Ca^{2+}}\)通道结合,引起\(\ce{Ca^{2+}}\)流入细胞质基质,通过与钙调蛋白结合引发进一步的细胞反应。
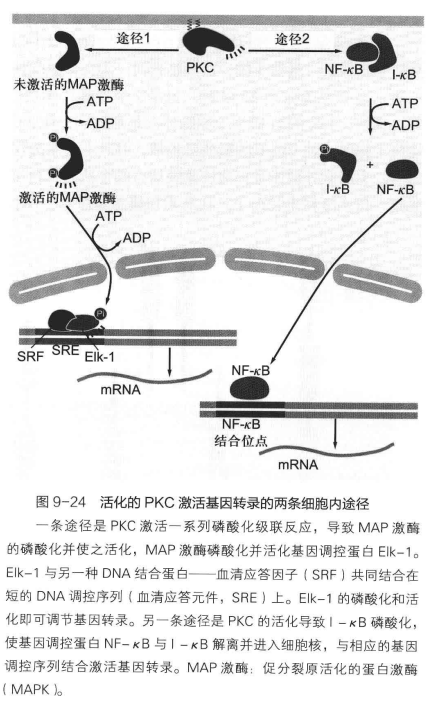
DAG 也没闲着,其结合在质膜上,负责参与活化质膜上的PKC(也就是蛋白激酶C)。特别地,静息状态下 PKC 一般在细胞质基质中,但是 IP3 使得内质网释放\(\ce{Ca^{2+}}\),细胞质基质中钙离子浓度上升,PKC 与之结合,转位到质膜上,在磷脂酰丝氨酸的共同作用下被 DAG 活化。其也是一种丝氨酸/苏氨酸蛋白激酶,通过磷酸化多种蛋白来调节其活性,进而影响细胞的状态。
两者可谓是天作之合,一个向内打开钙离子通道,一个在膜上活化 PKC,共同完成了对信号的接力转导。
钙离子/钙调蛋白信使体系
前面我们说了 IP3 介导的钙离子释放,特别地,\(\ce{Ca^{2+}}\)也是第二信使的一员,其主要通过浓度的升高与降低来传递信息。
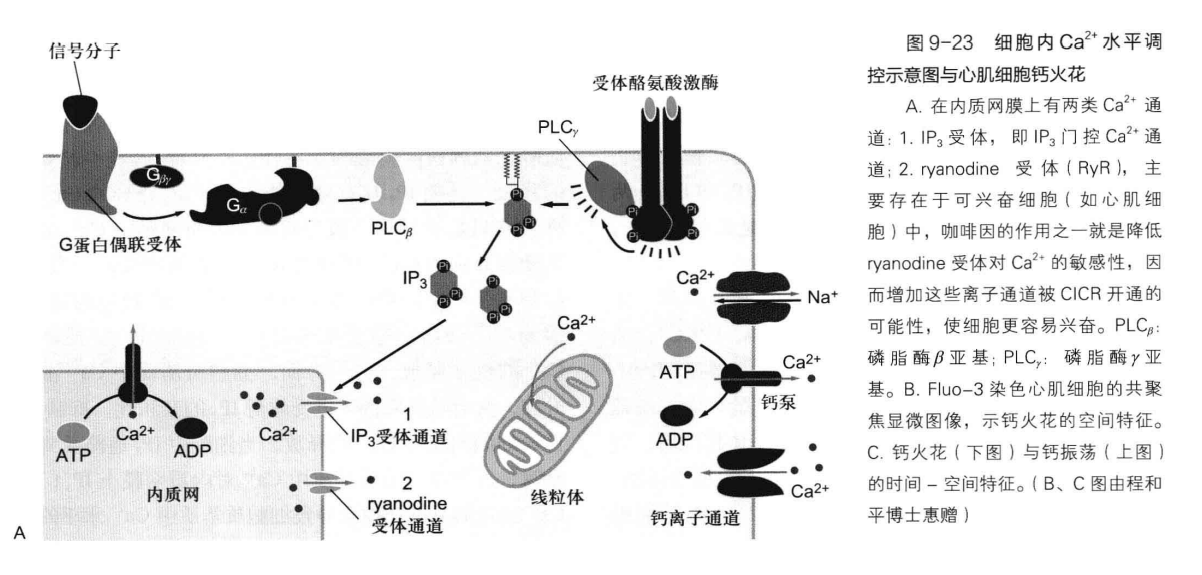
介导\(\ce{Ca^{2+}}\)信号转导的三种主要离子通道为:
- 质膜上电压依赖\(\ce{Ca^{2+}}\)通道
- ER上 IP3 门控\(\ce{Ca^{2+}}\)通道
- ryanodine 受体\(\ce{Ca^{2+}}\)通道
下图展示了一个经典的细胞内\(\ce{Ca^{2+}}\)水平调控:
钙离子作用机制
一般情况下\(\ce{Ca^{2+}}\)不会亲自去调控靶蛋白,而是通过\(\ce{Ca^{2+}}\)应答蛋白(或者说\(\ce{Ca^{2+}}\)结合蛋白)来间接发挥作用。钙调蛋白 (calmodulin, CaM) 就是其中最出名,分布最广泛的一种。
CaM 由一条多肽链折叠而成,呈现哑铃构象,有四个结构域,可以结合四个\(\ce{Ca^{2+}}\),活化形成\(\ce{Ca^{2+}-CaM}\)复合物后可以与靶蛋白结合来活化它们。
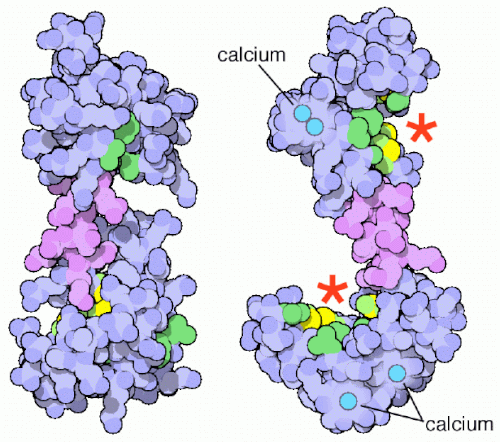
基于此原理,\(\ce{Ca^{2+}}\)可以有三条通路供它发挥:
- 信号刺激ER上\(\ce{Ca^{2+}}\)通道,释放内质网/肌质网中的\(\ce{Ca^{2+}}\)至细胞质基质中,激活 CaM 形成 \(\ce{Ca^{2+}-CaM}\)复合物,与靶蛋白结合并激活它们 (主要包括PhK、MLCK以及CaM-PK三种类型),从而激发细胞的反应。
- \(\ce{Ca^{2+}-CaM}\) 本身还可以通过激活细胞膜上的\(\ce{Ca^{2+}}\)泵,调节细胞内的\(\ce{Ca^{2+}}\)浓度。
- \(\ce{Ca^{2+}}\)也可直接对离子通道进行调节。
PI(3,4)P2 和 PI(3,4,5)P3 信使体系
PI(3,4)P2 和 PI(3,4,5)P3 由 PI 在磷脂酰肌醇-3-激酶 (PI3 - kinase, PI3K) 的催化下生成。特异的肌醇磷脂磷酸酶可使 PI(3, 4)P2 和PI(3,4,5)P3 去除3位磷酸。
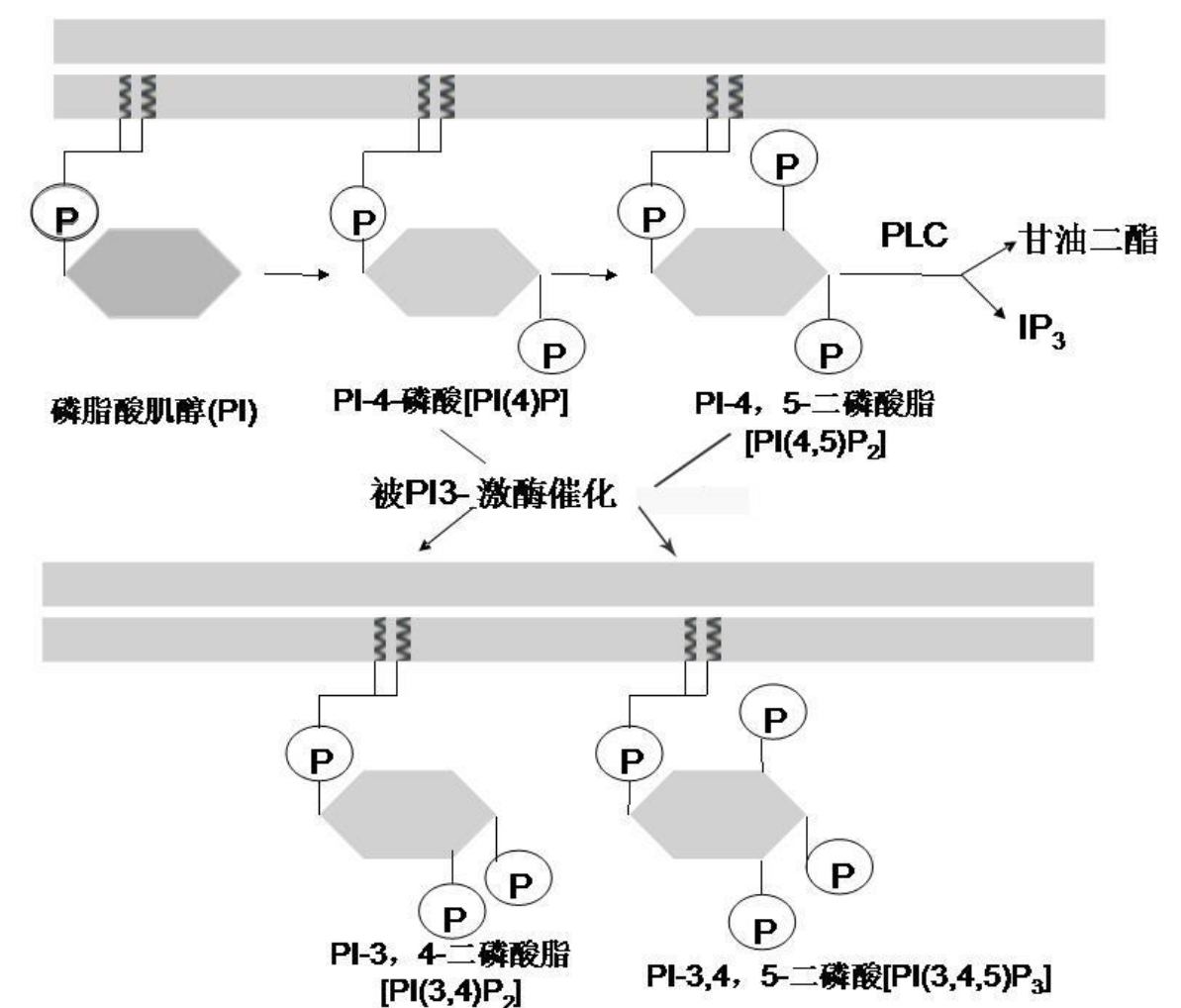
PI(3,4)P2 和 PI(3,4,5)P3 为多种胞内信号蛋白充当停泊位点,与信号蛋白的结合本身就是对信号蛋白的激活,进而介导多种下游信号通路。信号蛋白的停泊本身依靠其具有的 PH 结构域与其结合,形成信号传递复合体,将来自胞外的信号传递至胞内,促进细胞存活或生长。
信号转导与蛋白分子开关
蛋白分子开关
大部分胞内信号蛋白的功能上就像一个分子开关 (molecuar switch):当接受信号后信号蛋白从无活性状态转换成激活状态,反之则关闭信号蛋白。根据作用方式不同,分子开关分为两大类:GTPase分子开关和磷酸化激活/失活的蛋白质分子开关。
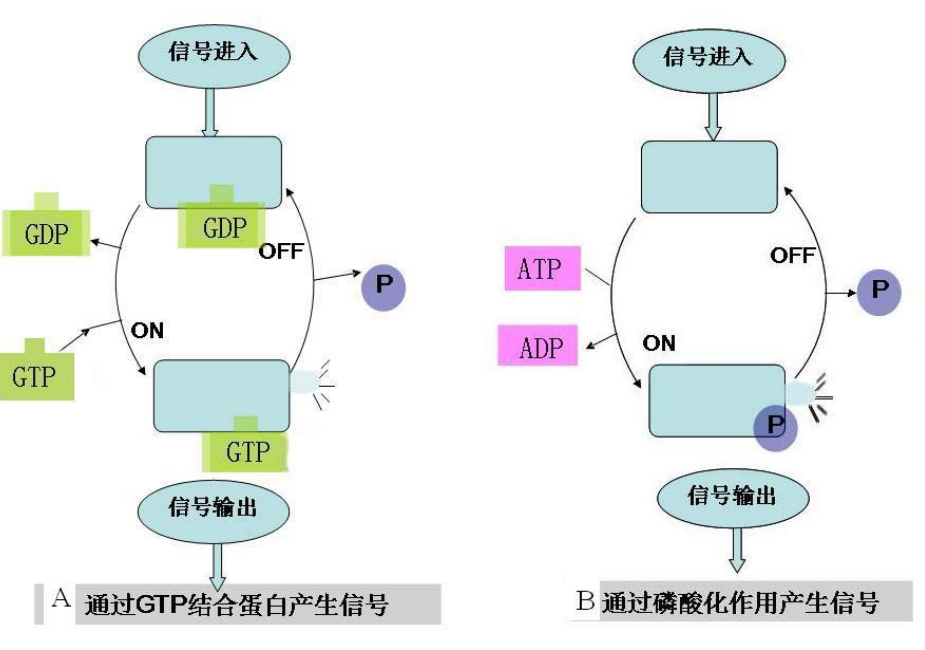
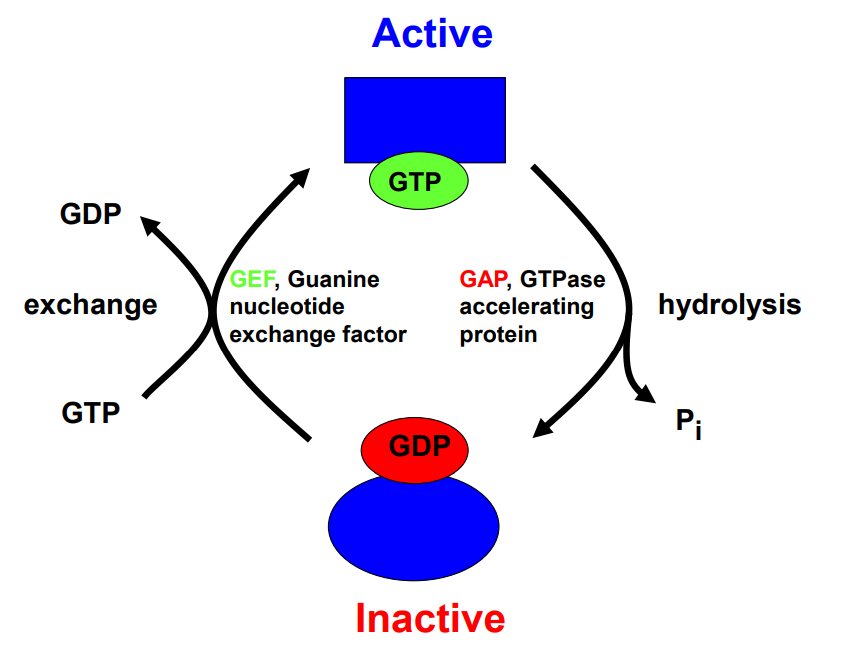
GTPase分子开关
GTPaser,也称为GTP 结合蛋白 (GTP binding protein)。在细胞内与信号转导相关的 GTPase 超家族包括:
- 三聚体GTP结合蛋白,典型的比如G蛋白
- 单体GTP结合蛋白,比如Ras蛋白与类Ras蛋白
当一个 GTPase 结合 GTP 时候被激活,GTP 水解为 GDP 时候被抑制。
磷酸化激活/失活分子开关
部分信号蛋白的状态由其是否被磷酸化所决定,蛋白激酶会对信号蛋白进行磷酸化修饰,就像是拨动了一个开关,使得信号蛋白构象变化,被激活/抑制。
而蛋白磷酸酶则是催化去磷酸化,与蛋白激酶作用相反。
根据作用底物氨基酸残基的特异性,蛋白激酶可以分为:蛋白质酪氨酸激酶与蛋白质丝氨酸/苏氨酸激酶。
蛋白质酪氨酸激酶,也就是 PTK,参与细胞生长、增殖与分化。主要分为膜上受体PTK与胞质非受体PTK,前者就包括了受体酪氨酸激酶 (RTK)。
丝氨酸/苏氨酸激酶,也就是 STK,也是体内大部分蛋白激酶所属的类别。活化后催化丝氨酸/苏氨酸残基的磷酸化。包括了 PKA (cAMP激活), PKC (DAG激活,钙库钥匙),PKG (cGMP激活),钙调蛋白依赖性蛋白激酶 (CaMK) 和丝裂原激活激酶 (MAPK)等。
磷酸级联反应
在细胞信号转导的过程中,许多胞内信号分子自身就是蛋白激酶,而它本身又可被上游的蛋白激酶磷酸化而激活,由此引起细胞内一系列蛋白质的磷酸化,产生级联效应(cascade),胞外信号分子所产生的信号被逐渐放大,在短时间内引起细胞效应。
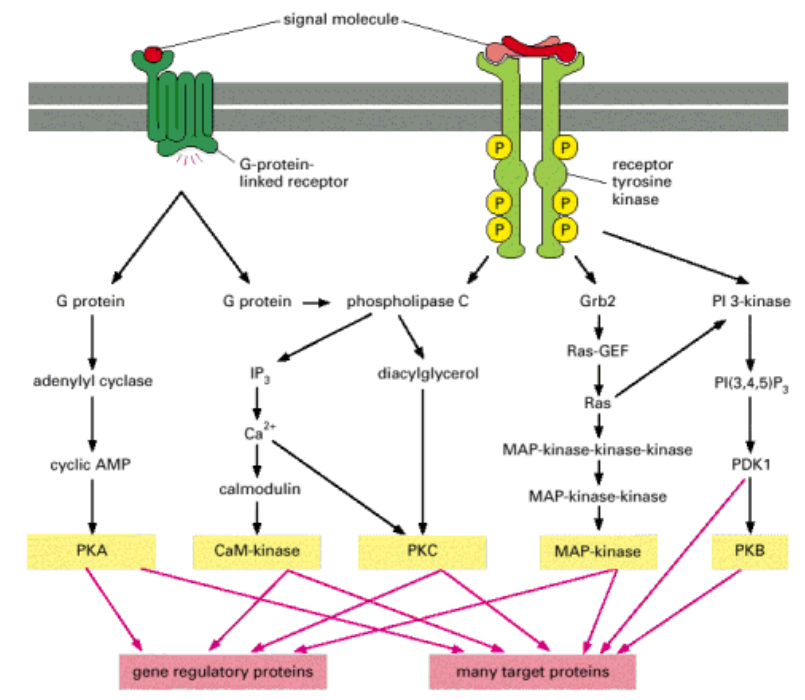
信号通路
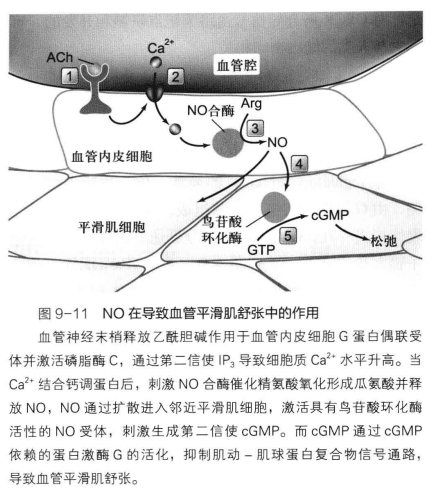
NO 信号通路
NO 是一种具有自由基性质的脂溶性气体分子,上游信号转导(比如乙酰胆碱)被血管上皮细胞受体捕获,激活 PLC,提升胞内钙浓度,激活 NO 合成酶催化精氨酸生成 NO。NO通0过扩散至邻近平滑肌细胞激活 GC 产生大量 cGMP,从而激活 PKG,使血管平滑肌舒张放松。
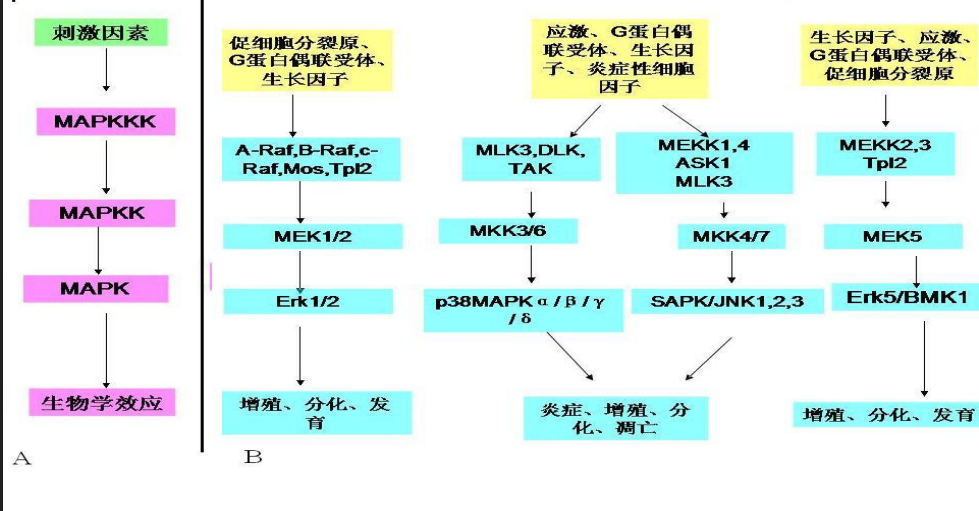
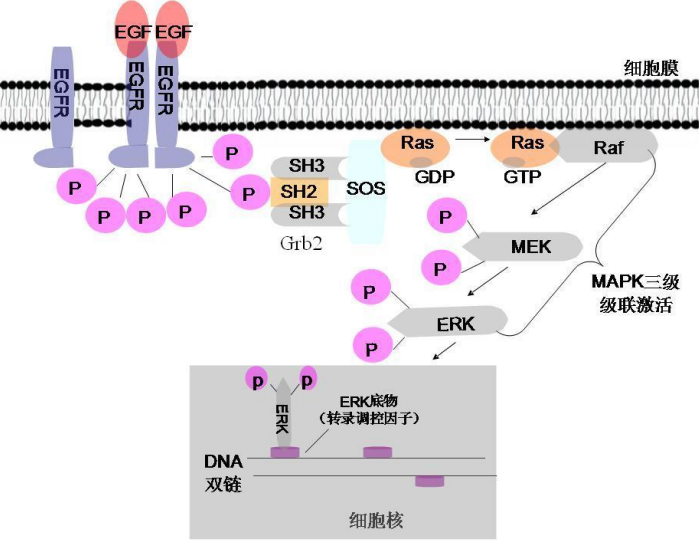
MAPK 信号通路
一个比较经典的信号通路如下,首先一个 RTK (比如图中的 EGFR),其通过具有SH2、SH3双结构域的接头蛋白链接下游的Sos(Ras-鸟苷酸交换因子),其与 Ras 结合,使得 Ras 上的 GDP 被交换为 GTP,使得其被激活,从而触发 MAPK 磷酸化级联反应。
Raf作为MAPKKK最先被Ras激活,作为丝氨酸/苏氨酸激酶活化作为MAPKK的 Mek,Mek 也作为蛋白激酶活化下游作为 MAPK 的 Erk,后者进一步传到具体的靶蛋白上。
JAK-STAT 信号通路
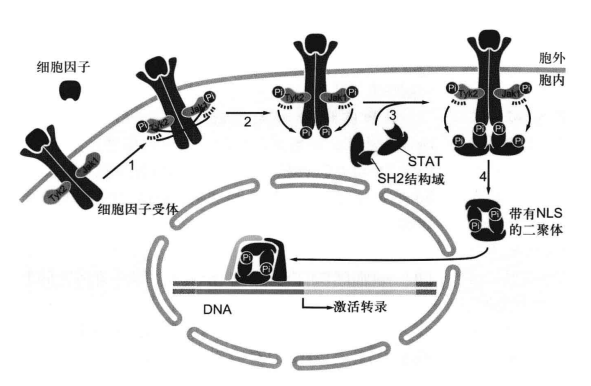
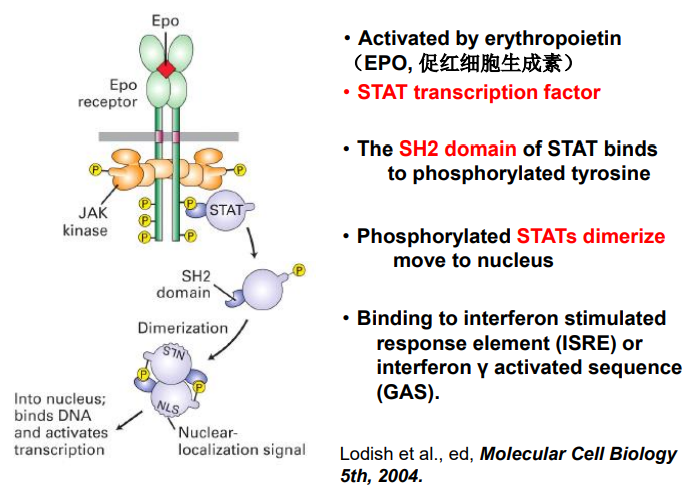
JAK 是一种非受体型酪氨酸激酶,需要酪氨酸激酶关联受体来激活它。比较经典的,与之相关的酪氨酸激酶关联受体是细胞因子受体,其感受 IL, IFN之后招募 JAK 并活化它们。JAK会将具有 SH2 结构域的 STAT 磷酸化酪氨酸残基。STAT 激活后从受体脱离,两两二聚化暴露 NLS,入核调控基因。
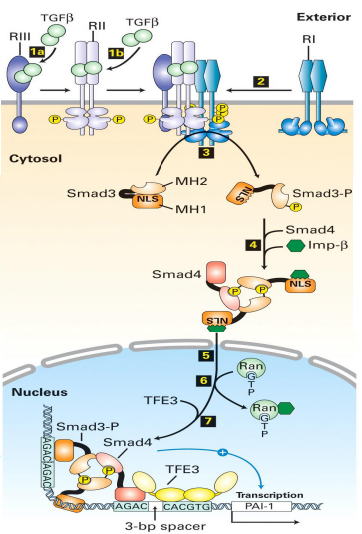
TGF-β 信号通路
TGF-β 信号通路,也就是转化生长因子β的信号通路,是对细胞周期具有强烈的抑制作用。
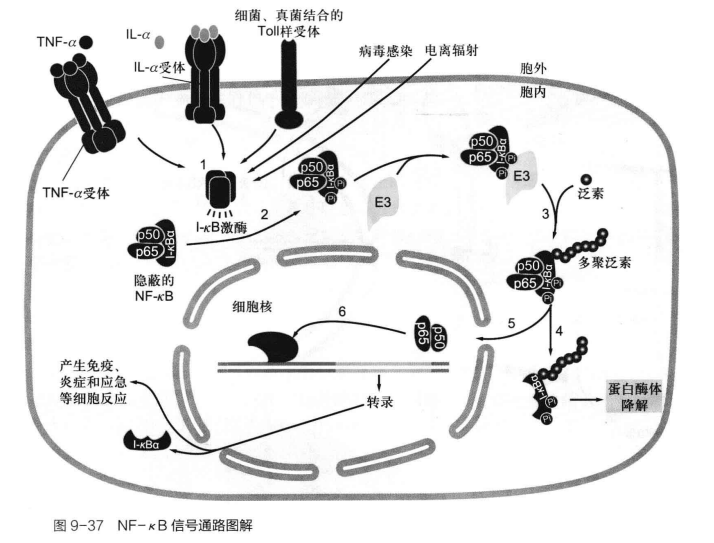
NF-κB 信号通路
这个信号通路与病原体入侵深刻相关,受体激活下游蛋I-κ激酶IKK磷酸化NF-κB抑制蛋白I-κB,使得后者降解,释放Nf-κB两个亚基p65,p50,暴露NLS入核,表达,CCL2,COX2,VEGF等一系促生长,趋化因子。
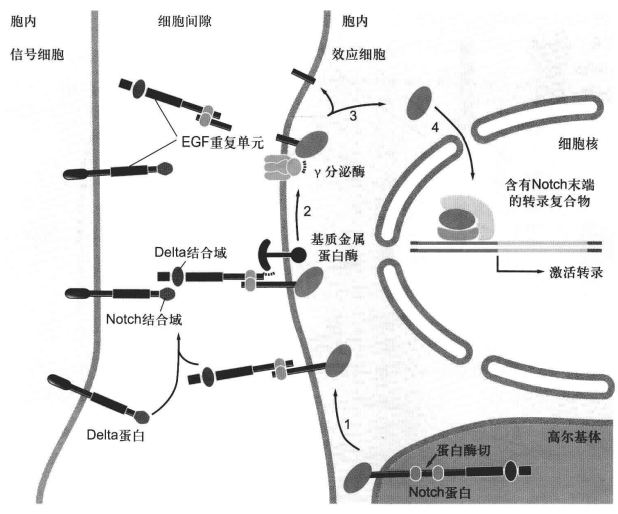
Notch 信号通路
该通路主要依赖于细胞间的直接接触,主要印象细胞的分化命运决定(胚胎发育过程中细胞间接触导致侧向分化抑制)。
Notch 信号通路还在细胞集群运动中发挥重要运动。
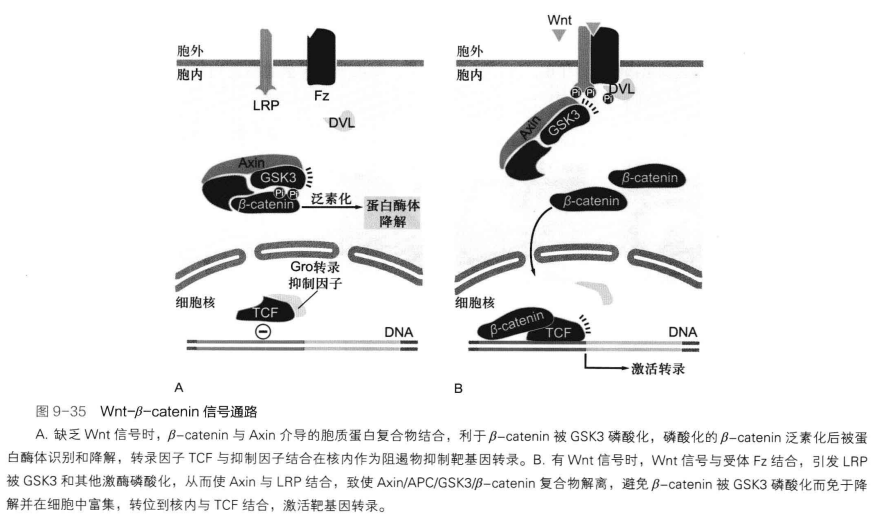
Wnt-β-catenin 信号通路
与维持细胞全能性有重要作用,由Wnt蛋白、相应受体及调节蛋白等成分构成。
信号转导的特点与网络调控
- 信号转导分子的激活机制具有类同性
- 信号转导过程为级联式反应
- 信号转导途径具有通用性与特异性
- 胞内信号转导途径相互交叉
- 整合作用