脂代谢
脂质的分类与结构
脂质是一类不溶于水而易溶于有机溶剂的化合物,在体内的脂质可以分为储能脂质和结构脂质。
- 储能脂质:包括脂肪酸、甘油酸酯,主要用于能量代谢和能量储存。
- 结构脂质:磷脂、糖脂、鞘脂、固醇等类脂,存在生物膜中。
甘油三酯
甘油三酯 (TG),又称为三酰甘油酸 (TAG),也是食物性脂肪的主要存在形式,在人体内主要存储在脂肪组织的脂滴中。
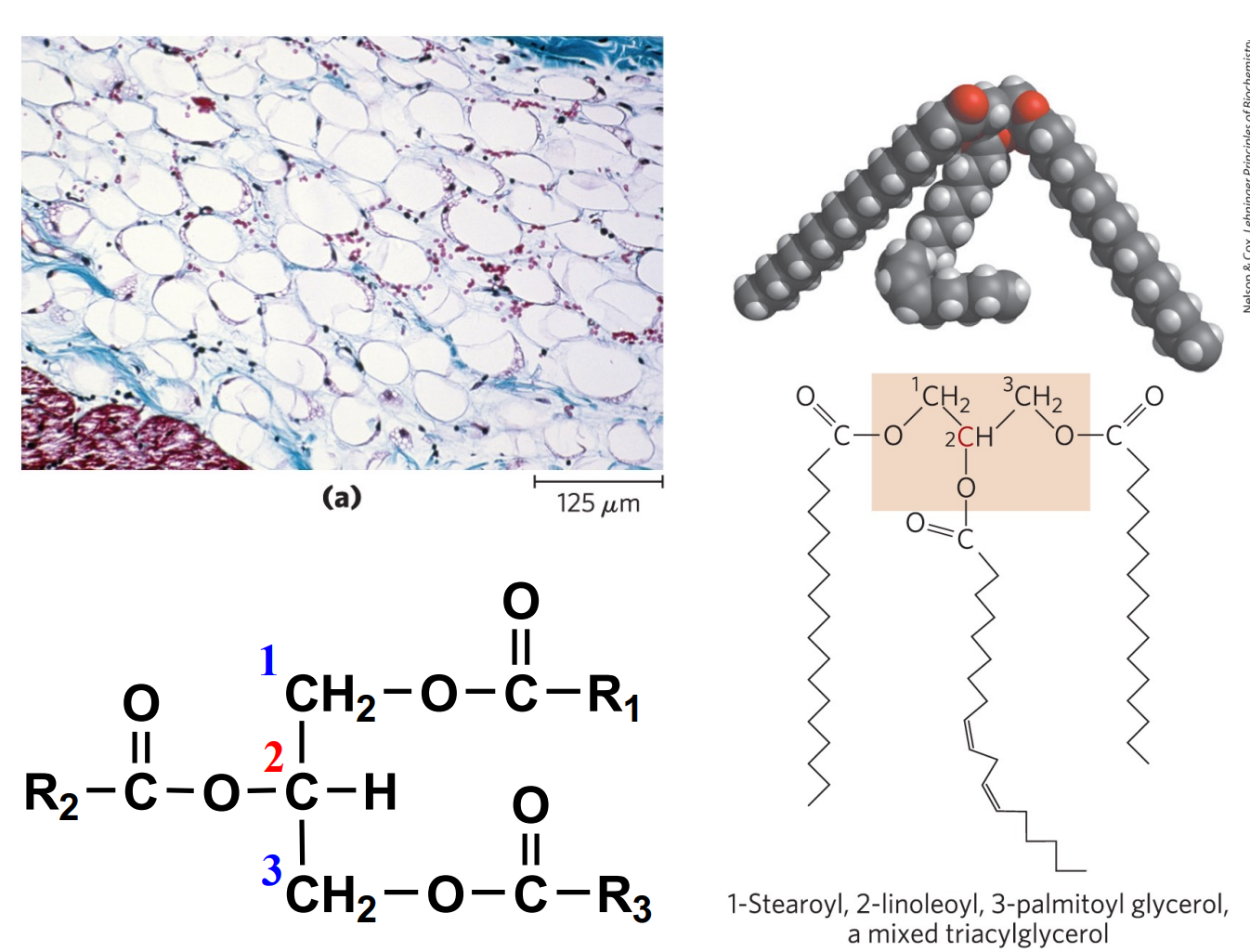
常见脂肪酸
脂肪酸根据是否能在人体内从头合成,分为非必需脂肪酸与必需脂肪酸:
- 非必需脂肪酸:机体能从头合成的,包括软脂酸(16C饱和脂,又称为棕榈酸),硬脂酸(18C饱和),棕榈油酸(16C单不饱和)和油酸(18C单不饱和)。
- 必需脂肪酸:机体必需的脂肪酸,但是又无法从头合成/合成量不足,一般从植物油中获取,包括亚油酸、亚麻酸和花生四烯酸。
Note
通常饱和脂肪酸的熔点高于非饱和脂肪酸
脂肪酸命名方式
- \(\Delta\)编码体系:从羧基碳原子起计双键位置。
- \(\omega\)或\(n\)编码体系:从甲基碳起计双键位置。
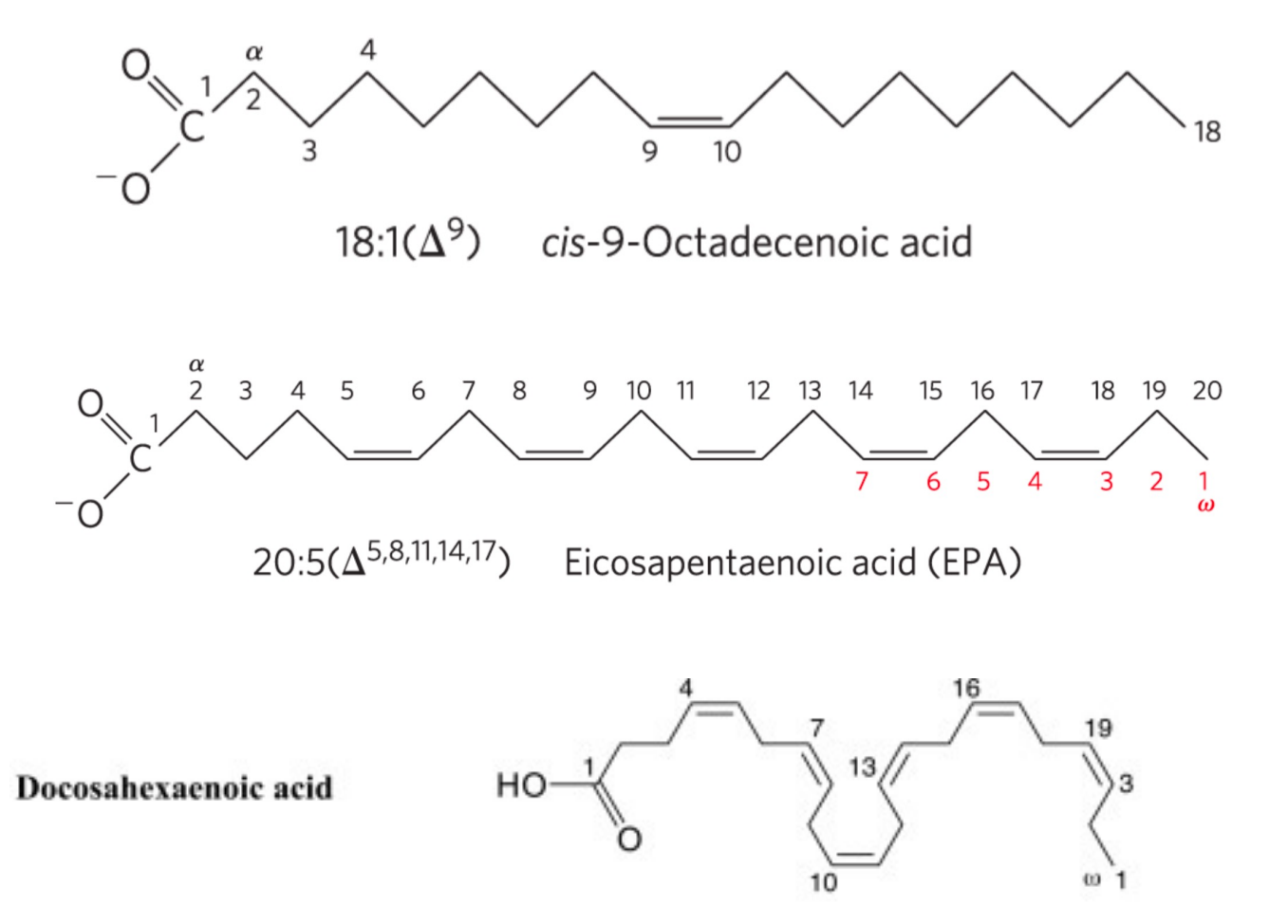
结构脂质分类
结构脂质又称为类脂,动物的生物膜中主要包含磷脂、糖脂和固醇,其中磷脂和固醇的含量较高。
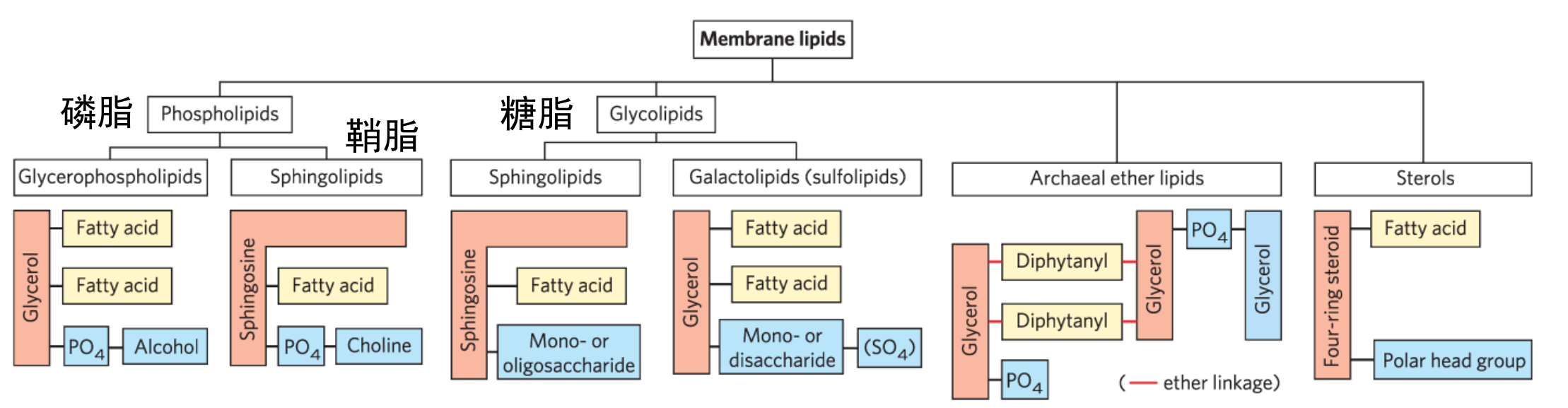
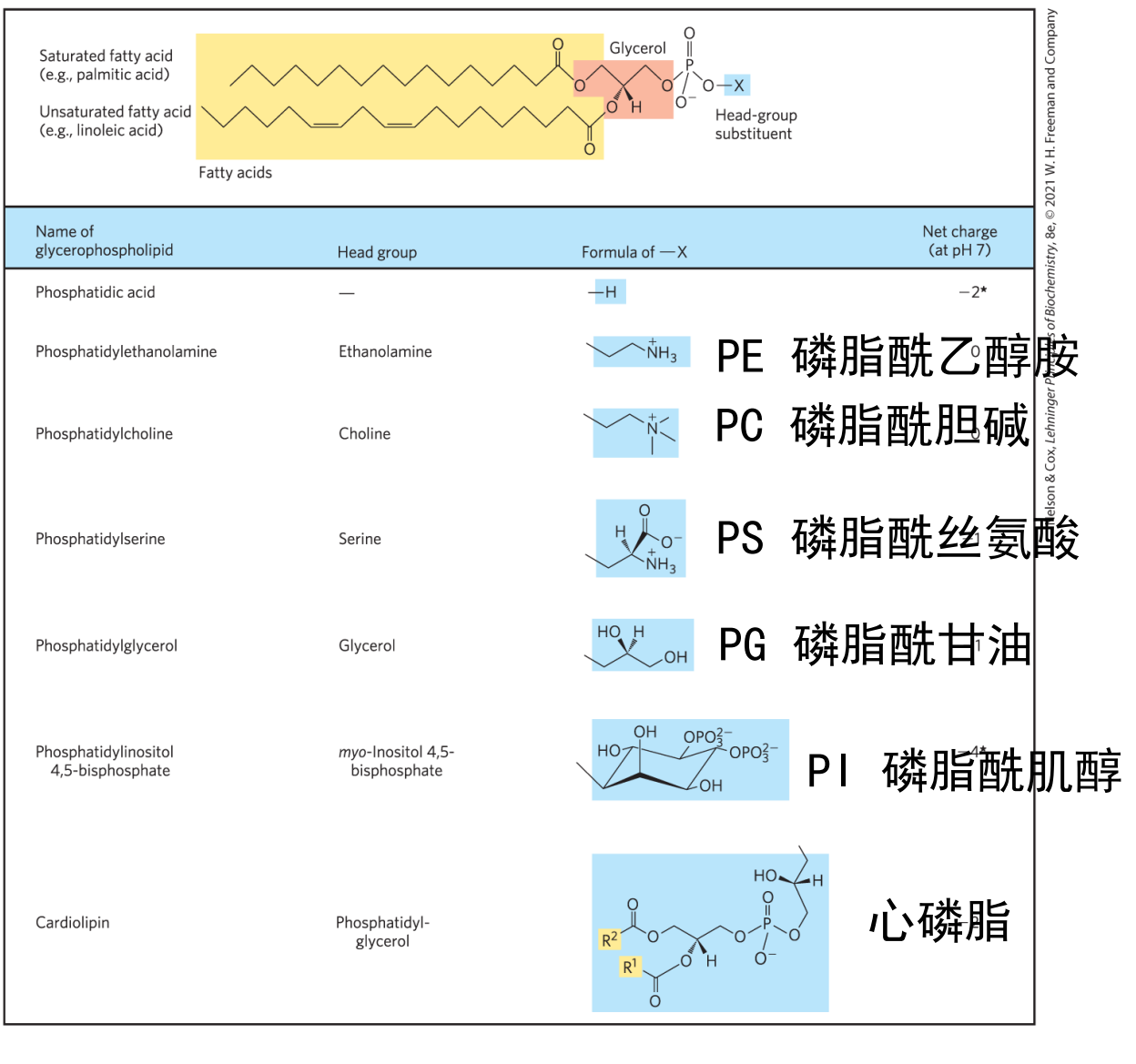
甘油磷脂
甘油磷脂是生物膜的主要组成部分,其在第一、二个碳上分别连接着一条脂肪酸链(一般情况饱和:不饱和 = 1:1),第三个碳上通过磷脂键连接有高度极性或者带电的基团。
根据磷酸基团头部的不同,可以将磷脂分为以下几类:
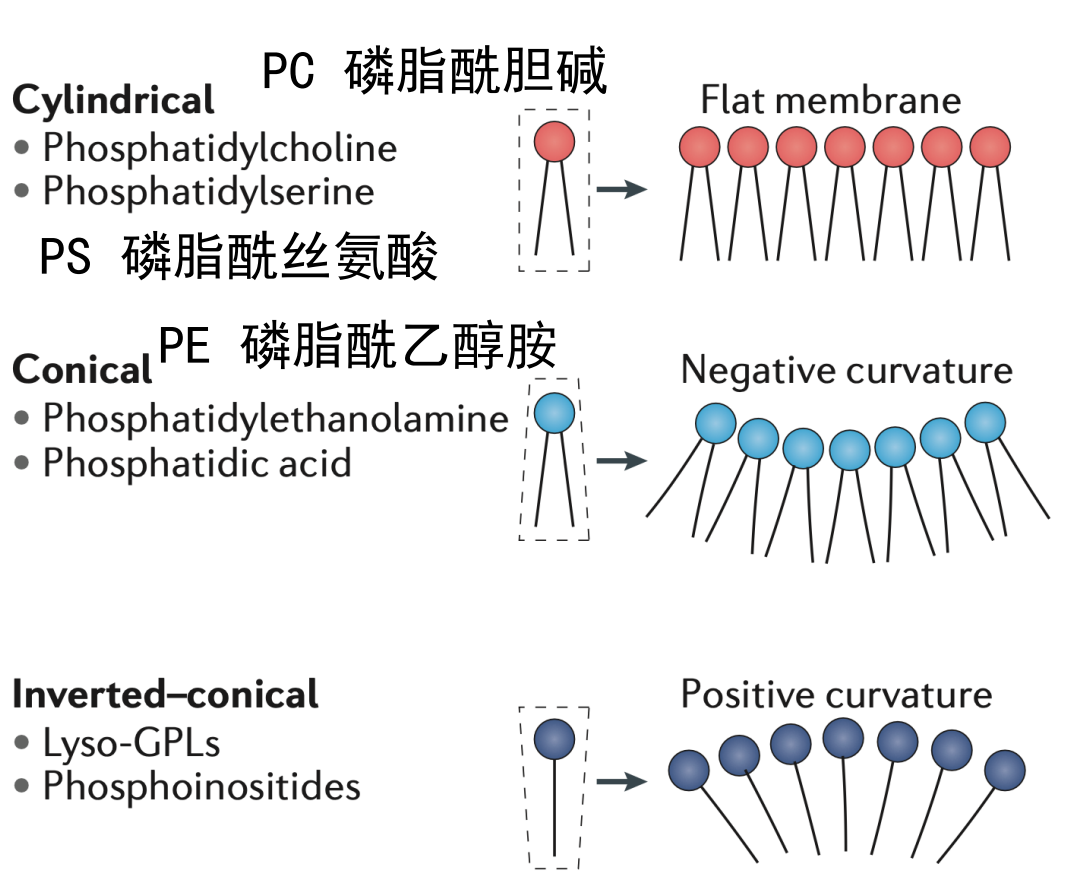
磷脂与膜曲率
不同的磷脂形成的膜结构会天然形成不同正负取向的膜曲率。
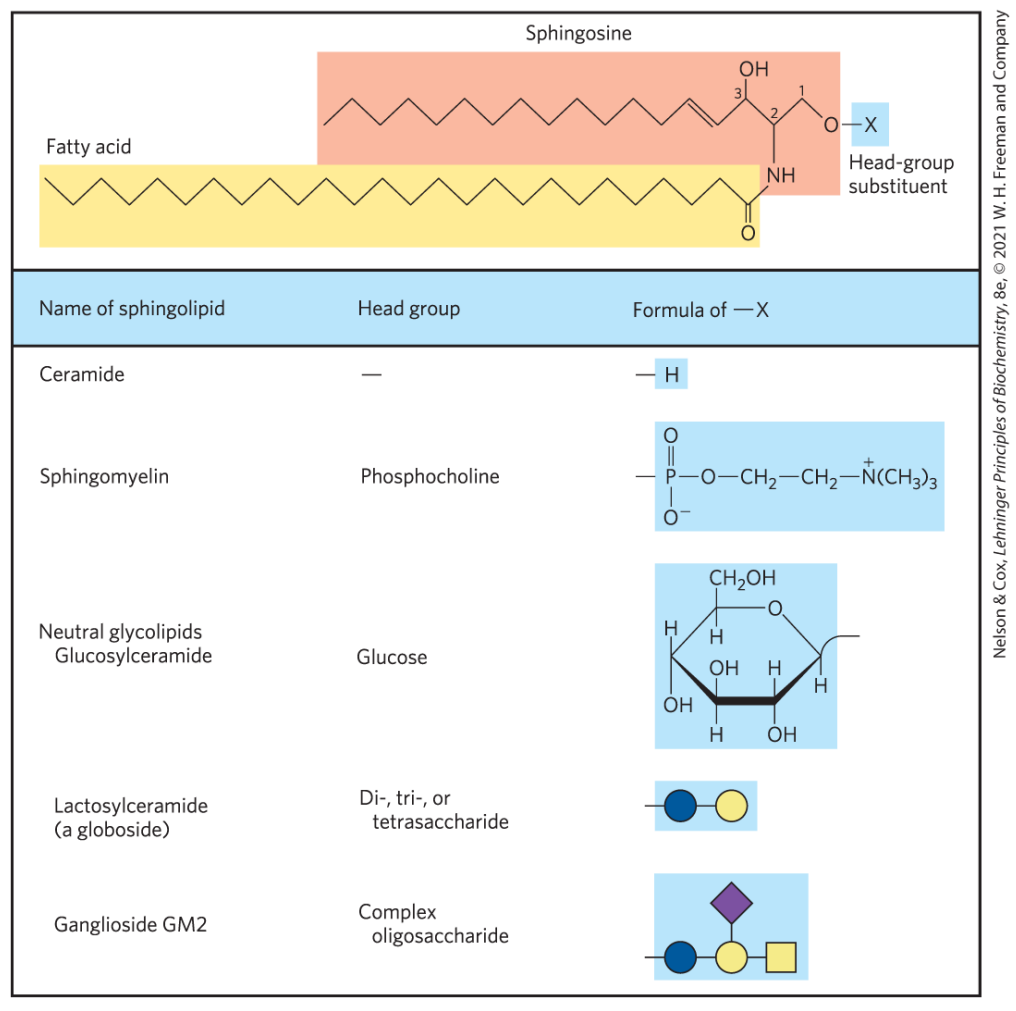
鞘脂
鞘脂是一类含有鞘氨醇或二氢鞘氨醇的脂类,根据头部取代基的不同又可以分为鞘磷脂和鞘糖脂。其在保护细胞膜与信号传导中发挥重要作用。
鞘脂的头部集团为氢原子时即为神经酰胺。
 、
、
固醇
固醇的最大特点是其具有四个碳环(通常还有一定刚性),故其也称为甾醇。动物细胞中含量最高的固醇是胆固醇,是调节膜流动性并充当激素的功能。
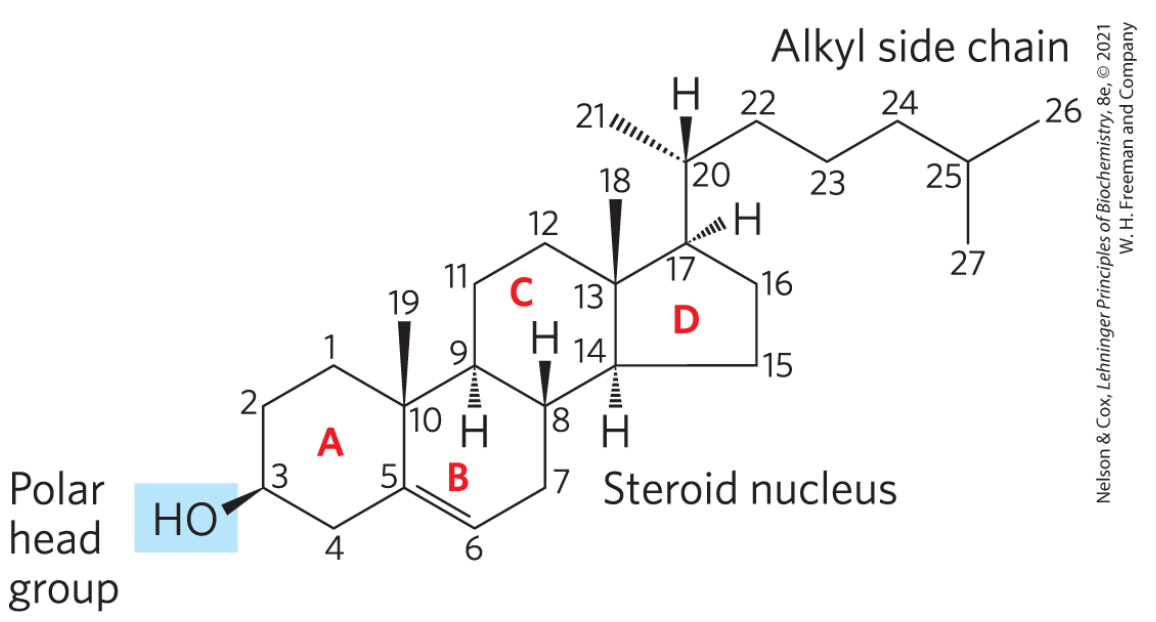
甾
“甾”其实是一个新造象形字,上面的三个撇点代表三个取代基,下面一个田代表四个碳环。
脂类消化与吸收
自进入消化系统中开始,胆囊释放的胆汁酸盐会将脂肪颗粒乳化为混合微团。
接着肠道中的脂酶水解甘油三酯等脂类,脂肪酸与其它降解产物进入小肠粘膜细胞被再次转化为甘油三酯。
甘油三酯、磷脂与胆固醇和载脂蛋白结合形成乳糜颗粒 (CM),被肠粘膜细胞分泌,经淋巴系统进入血液循环。
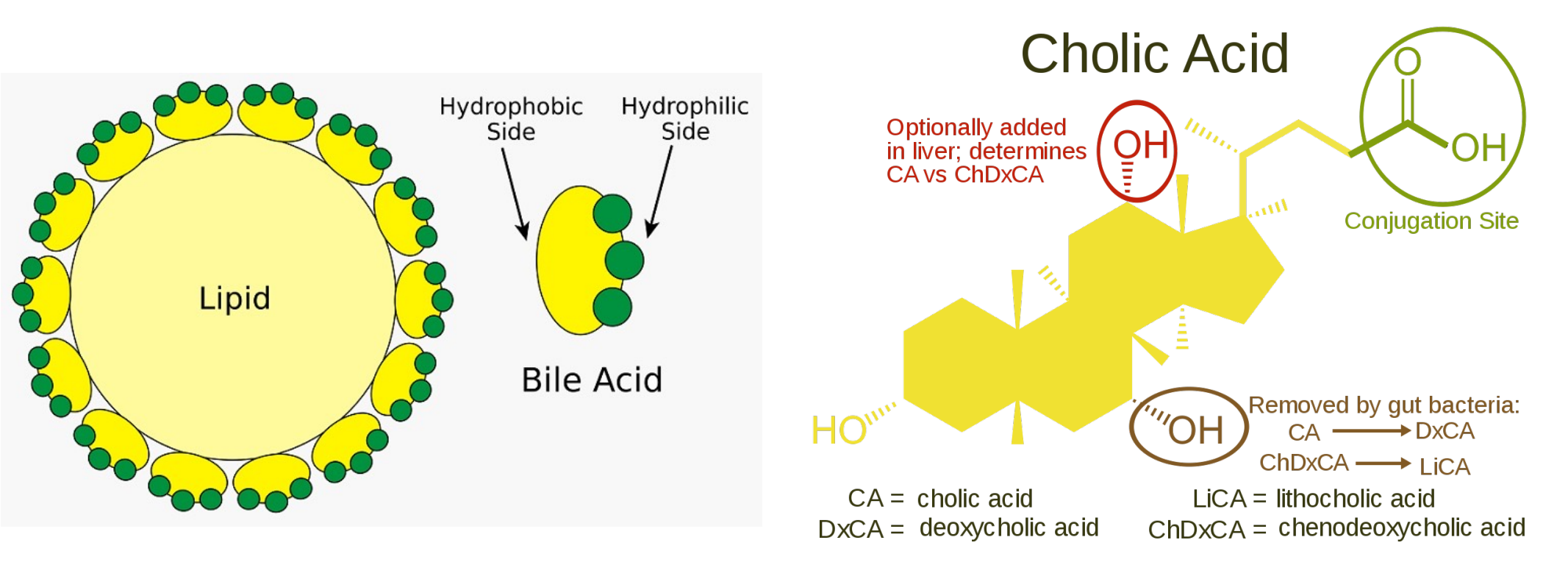
胆汁酸
胆汁酸类似于我们平日用的肥皂,是一种两性分子,可以乳化大滴脂肪,便于消化的进行。其会通过门静脉被回收至肝脏与胆囊。
脂酶
消化过程中主要参与的脂酶有以下几类:
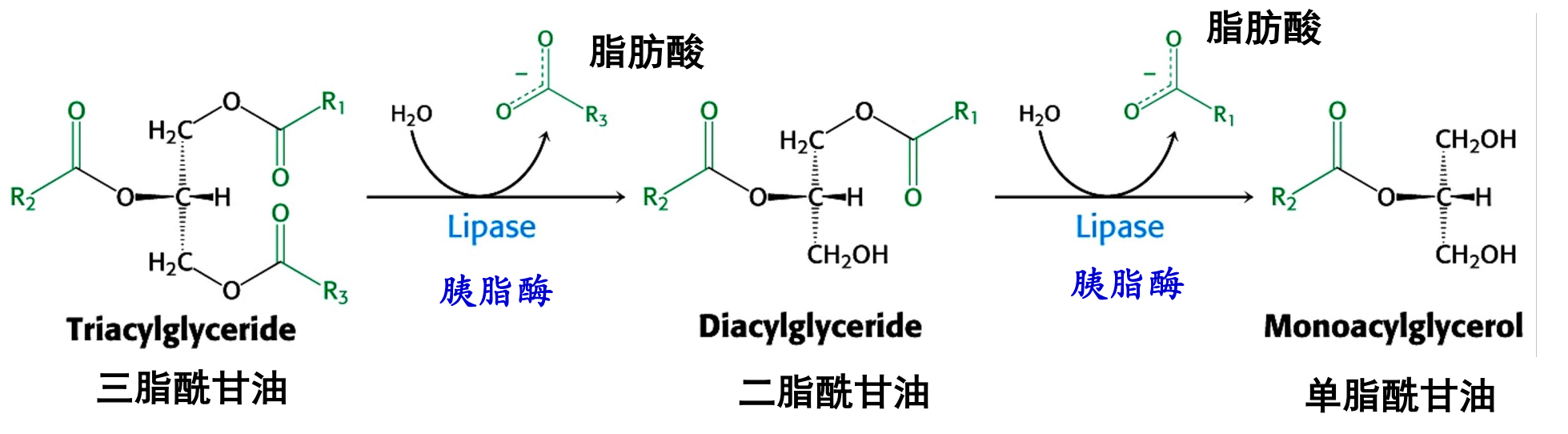
胰脂酶将甘油三酯水解为2-甘油一酯和脂肪酸。
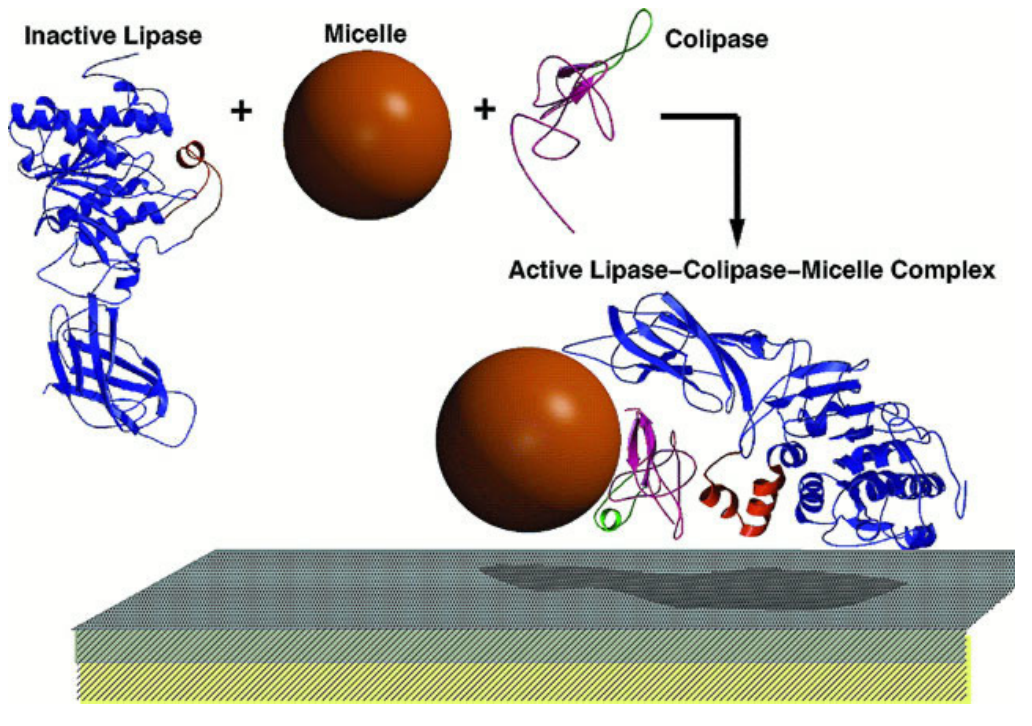
在过程中,辅脂酶协助磷脂酶发挥功能,其能与胰脂酶和胆汁酸盐结合,使得胰脂酶可以吸附在微团的水油界面上,有利于后者对甘油三酯的水解。
其次,辅脂酶还可以保护胰脂酶在界面上变性,解除胆汁酸盐对胰脂酶的抑制作用。
甘油三酯的分解代谢
脂肪动员
脂肪的水解受到激素的调控,包括有:
- 脂解激素:肾上腺素、胰高血糖素与去甲肾上腺素
- 抗脂解激素:胰岛素与前列腺素
对脂肪的动员过程实现脂肪细胞中脂肪的水解。
脂肪动员
储存在脂肪细胞中的脂肪被脂肪酶逐步水解为游离脂肪酸和甘油,并释放入血供其他组织氧化利用的过程,被称为脂肪动员。
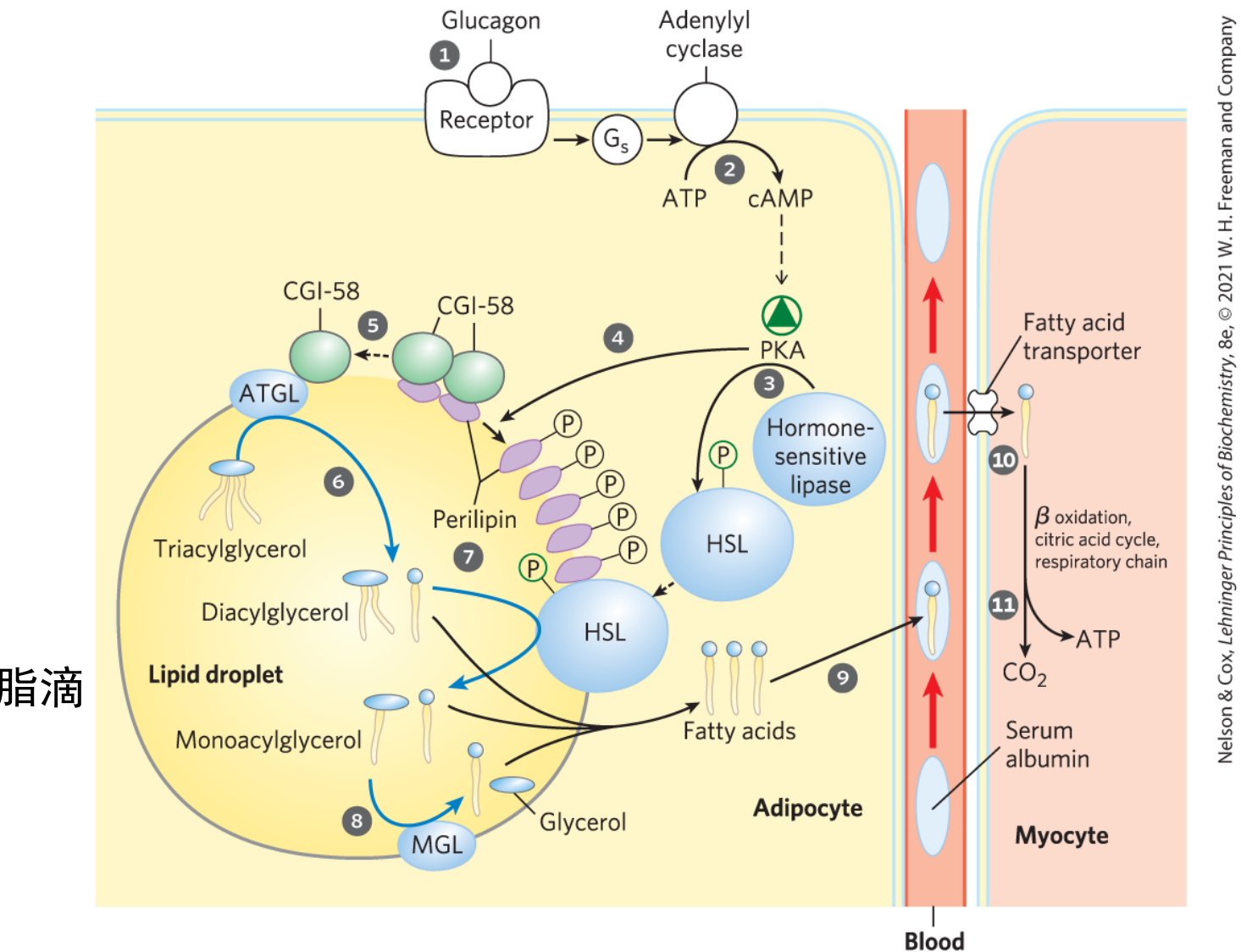
脂肪酶会依次参与甘油三酯的水解,次序为甘油三酯脂肪酶 (ATGL)、激素敏感性脂肪酶 (HSL)和甘油一酯脂肪酶 (MGL)。
脂肪动员后的去向
脂肪酸在血中主要由清蛋白运输,主要由心、肝和骨骼肌等摄取与利用。脂肪酸携带了95%脂肪的能量。
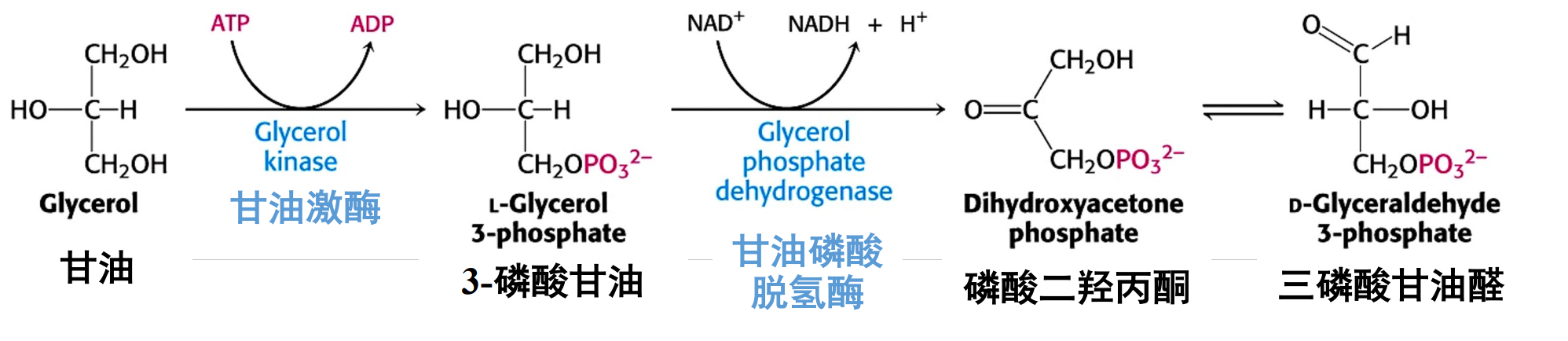
脂肪细胞不存在甘油激酶,骨骼肌等组织因甘油激酶活性很低,故不能很好利用甘油。其是甘油转换代谢中间物,从而进入糖代谢途径的关键酶,在肝脏中活性最高。
甘油直接运至肝、肾、肠等组织,主要在前两者进行糖异生。
清蛋白
血浆清蛋白约占血浆总蛋白量的50%,其上有7个脂肪酸的结合位点。脂肪酸通过与可溶于水的清蛋白结合,就可以通过血液在不同组织间转运。
脂肪酸的β-氧化
脂肪酸的β-氧化在肝、心肌与骨骼肌中最活跃,其过程主要包括:
- 脂酸活化——脂酰CoA的生成
- 脂酰CoA进入线粒体
- 脂酸的β-氧化
- 脂酸氧化的能量生成
脂酸活化——脂酰CoA的生成
该步骤在细胞质基质中进行,其在脂酰CoA合成酶的催化下生长脂酰CoA。
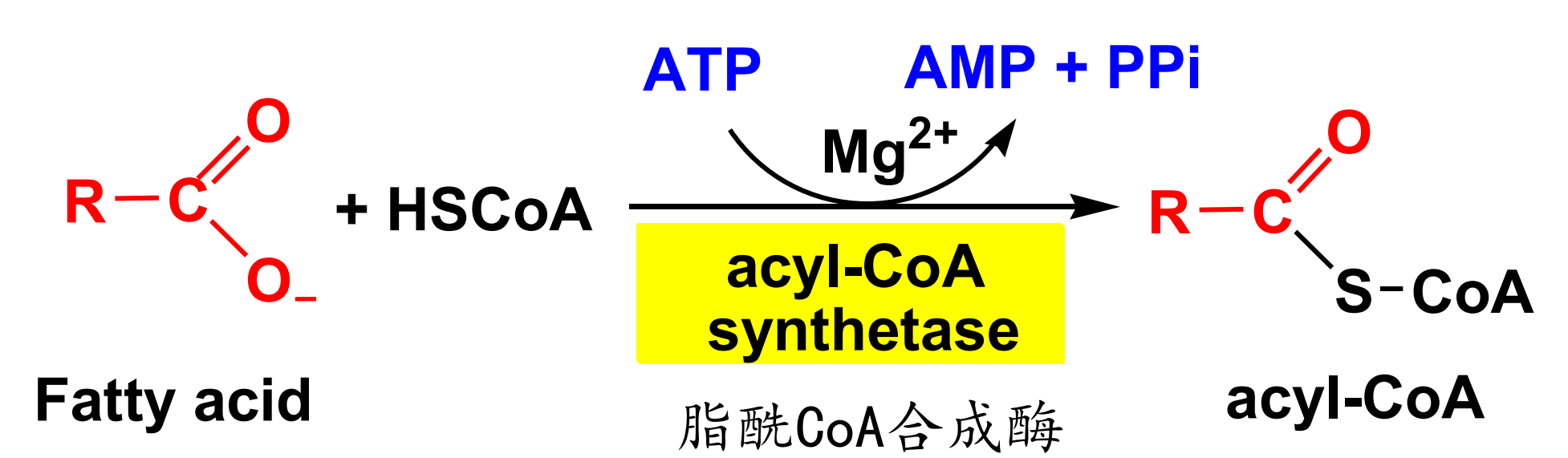
由于反应生成PPi,其会发生水解,故反应不可逆。
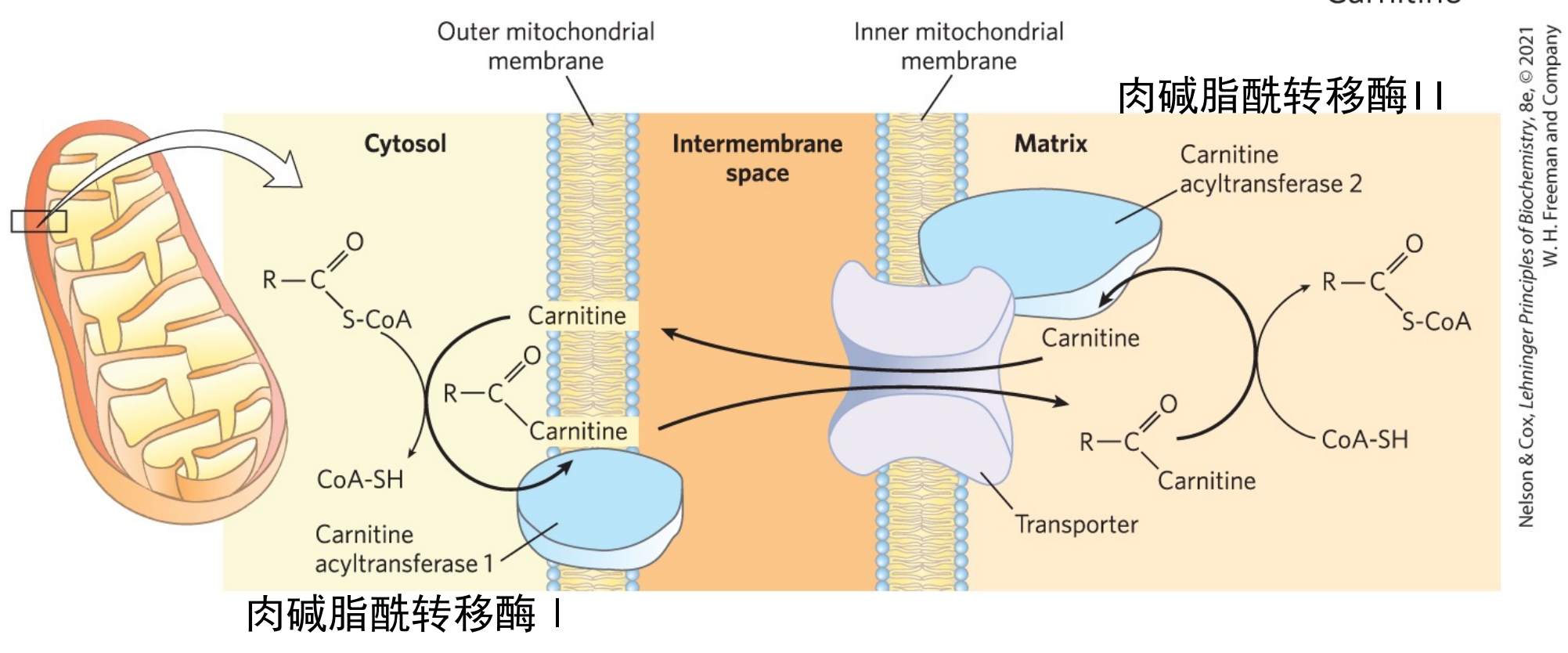
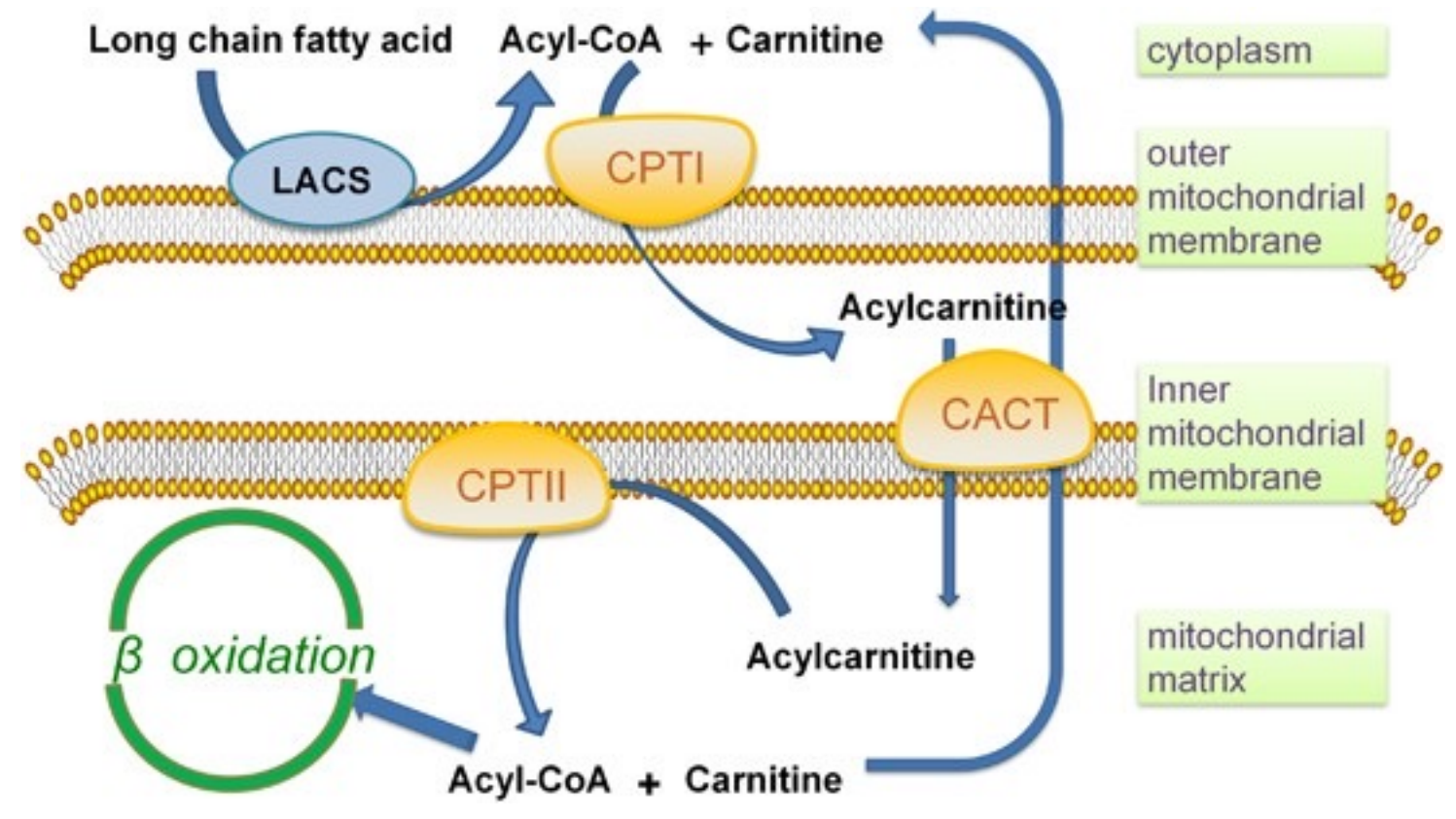
脂酰CoA进入线粒体
脂肪酸的活化在细胞质基质中进行,但是其进一步的氧化利用在线粒体基质中,所以活化生成的脂酰CoA必须在肉碱的帮助下进入线粒体内。
肉碱
脂酰CoA转运的调控
在转运过程中,肉碱脂酰转移酶I是限速酶,脂酰CoA进入线粒体是脂酸β-氧化的主要限速步骤。
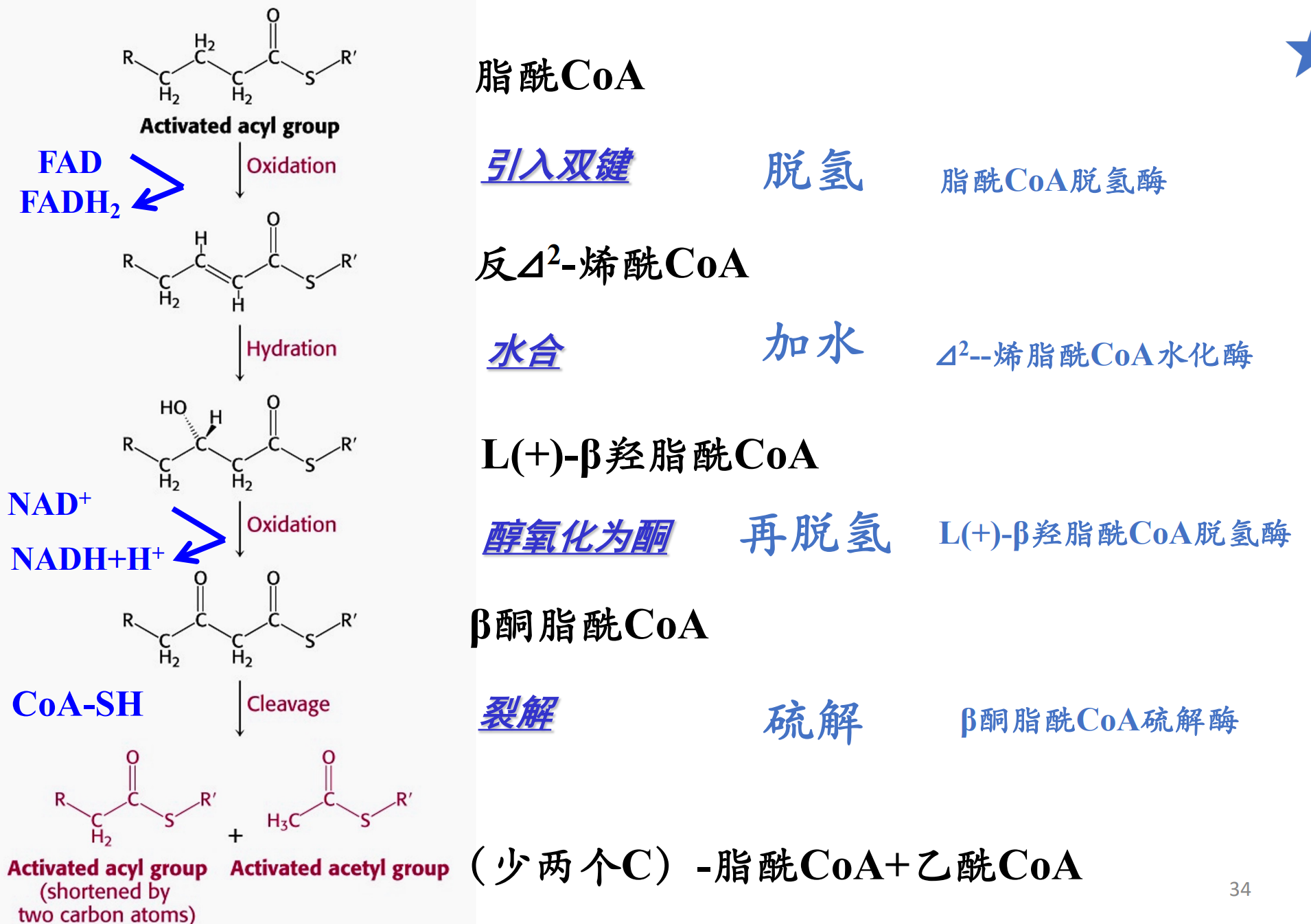
脂酸的β-氧化
在线粒体基质中,长链脂酰CoA会在脂肪酸氧化酶系的催化下发生β-氧化——从β-碳原子开始每次氧化断去二碳单位的乙酰CoA的过程。
生成的乙酰CoA可以通过TCA循环进一步释放能量。
偶数碳的脂肪酸可以完全生成乙酰CoA,而奇数碳的脂肪酸最终会生成乙酰CoA和一分子的丙酰CoA。
β-氧化
一个脂酸β-氧化循环包括:脱氢、加水、再脱氢与最后硫解,最终生成一分子乙酰CoA。
其中第一次脱氢由\(\ce{FAD}\)接受,第二次脱氢由\(\ce{NAD+}\)接受。
体内存在多种脂酰辅酶A,其具有不同的底物偏好,可以分为VLCAD、LCAD与MCAD。
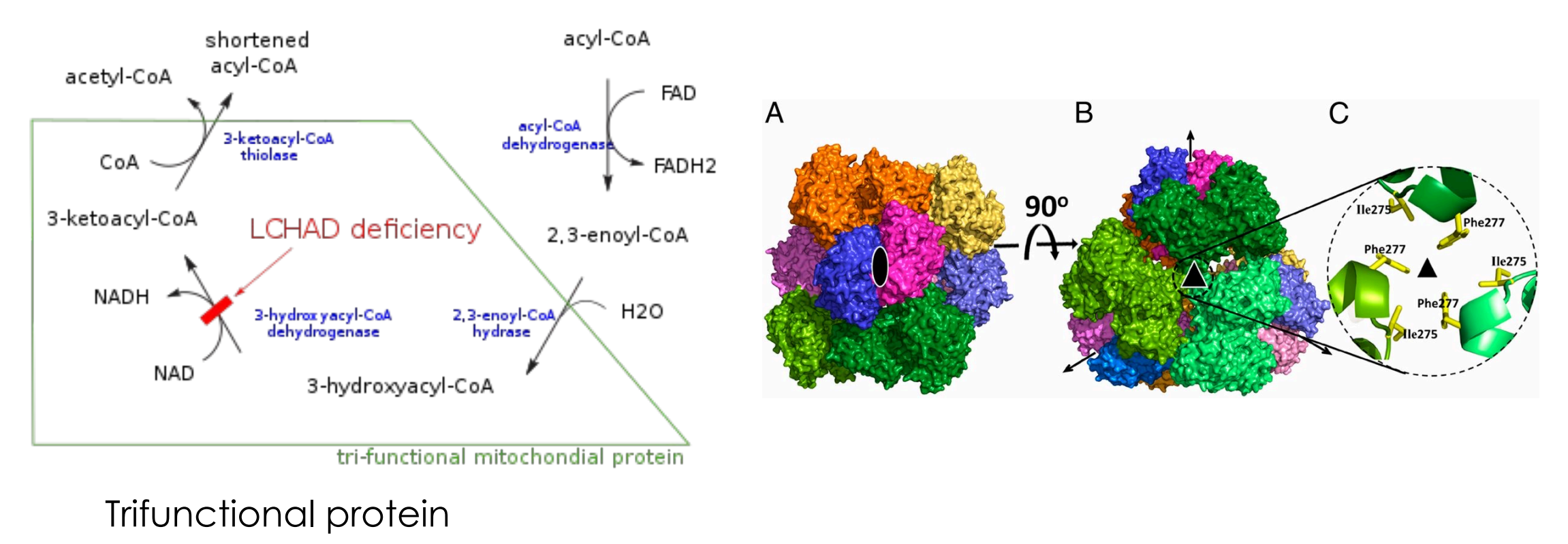
脂酸β-氧化的后三步同时由一个三重功能蛋白催化。
脂肪酸β-氧化的能量生成
需要强调的是:脂肪酸的β-氧化本身不能生成任何能量,只能生成乙酰CoA和供氢体,其要进一步释放能量需要借助TCA循环和氧化磷酸化。
脂肪酸的功能效率是葡萄糖无法比拟的,以软脂酸(16C饱和)为例,其一共可以生成106分子的ATP。
脂肪酸的其他氧化途径
- 不饱和脂肪酸的降解还需要异构酶和还原酶的参与。0
- 过氧化物酶体也可以氧化脂肪酸
- 丙酰CoA可以参与进一步的氧化。
不饱和脂肪酸的氧化
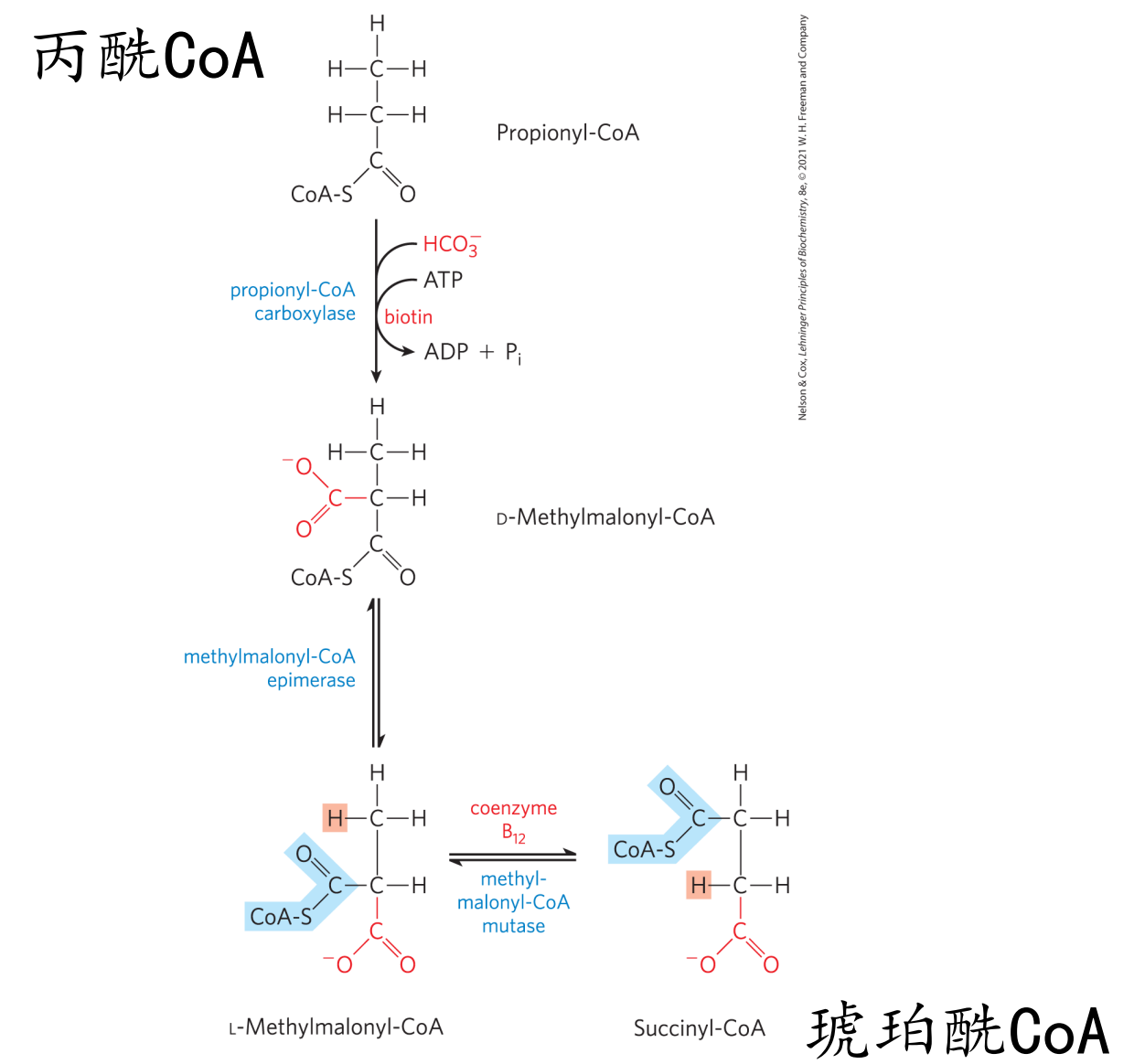
丙酰CoA的氧化
丙酰CoA在羧化酶和异构酶的作用下可以转变为琥珀酰CoA,再经三羧酸循环进行代谢。
奇数脂肪酸的部分骨架可以进入糖异生途径。
乙酰CoA的去路
- 进入TCA循环,最终氧化生成二氧化碳和水以及大量的ATP
- 在肝脏及肾脏细胞中生成酮体
酮体的合成与利用
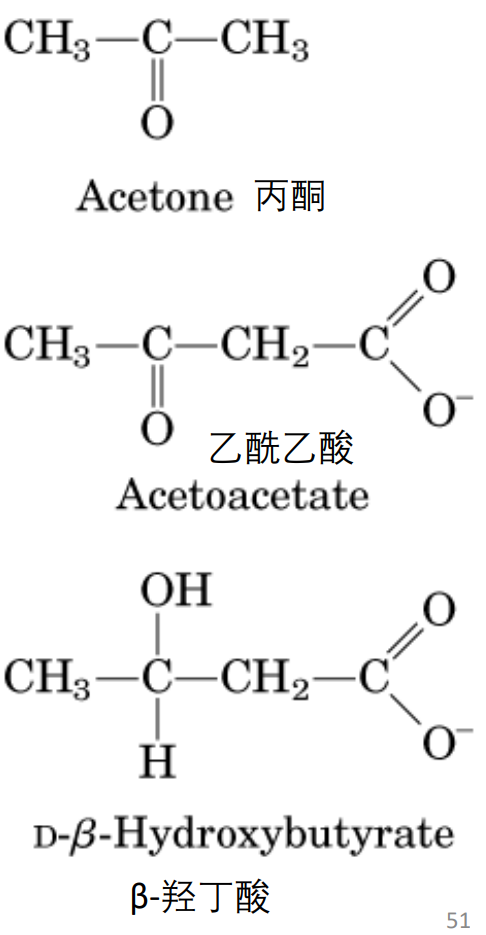
酮体是脂酸在肝分解氧化时特有的中间代谢产物,是乙酰乙酸、β-羟丁酸和丙酮三者的统称。
生酮饮食指的是低碳水高脂肪(碳水10%,脂肪70%)的饮食模式。
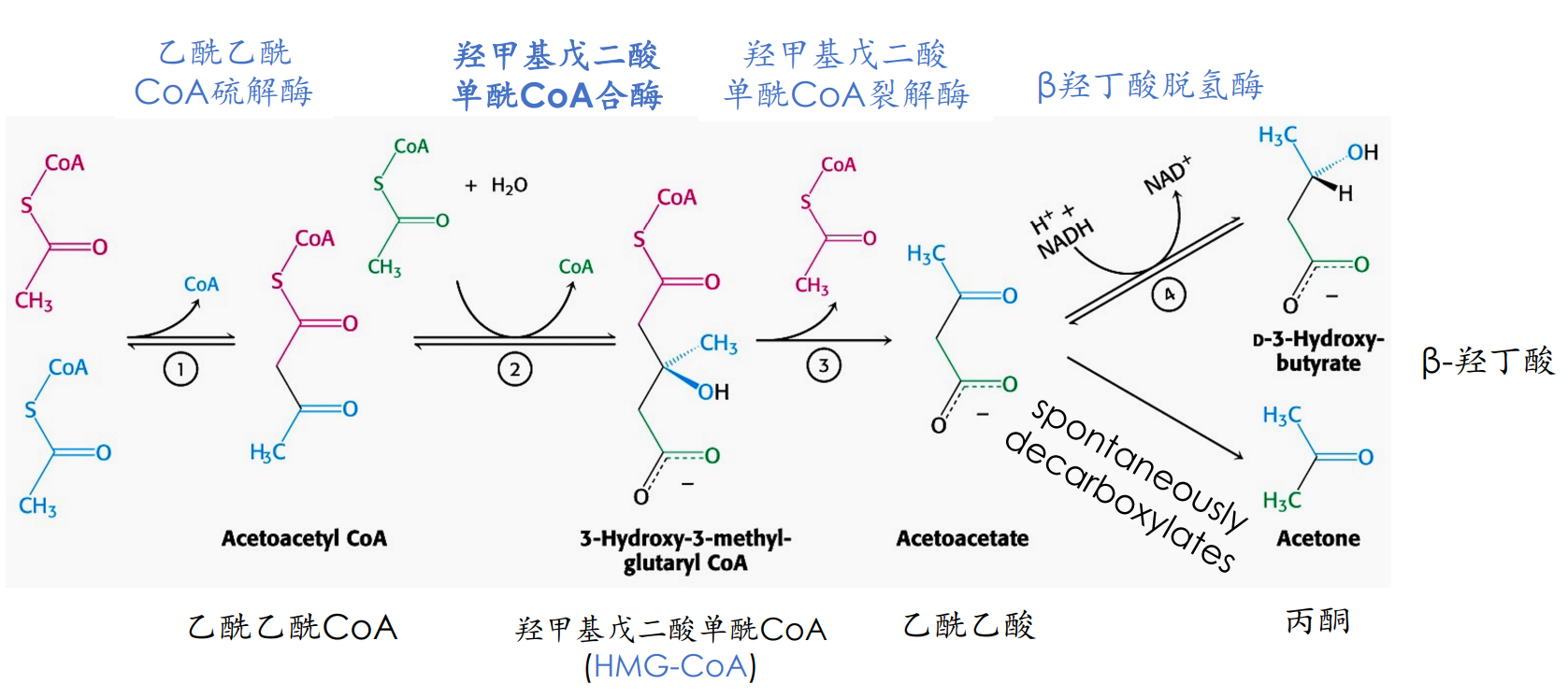
酮体的生成部位是肝线粒体,原料为乙酰CoA,主要来自脂肪的β-氧化,关键酶为HMG CoA合成酶。
酮体生成条件
- 草酰乙酸相对缺乏,乙酰CoA相对过量
- 禁食、饥饿或高脂肪饮食
- 糖代谢缺陷(糖尿病)
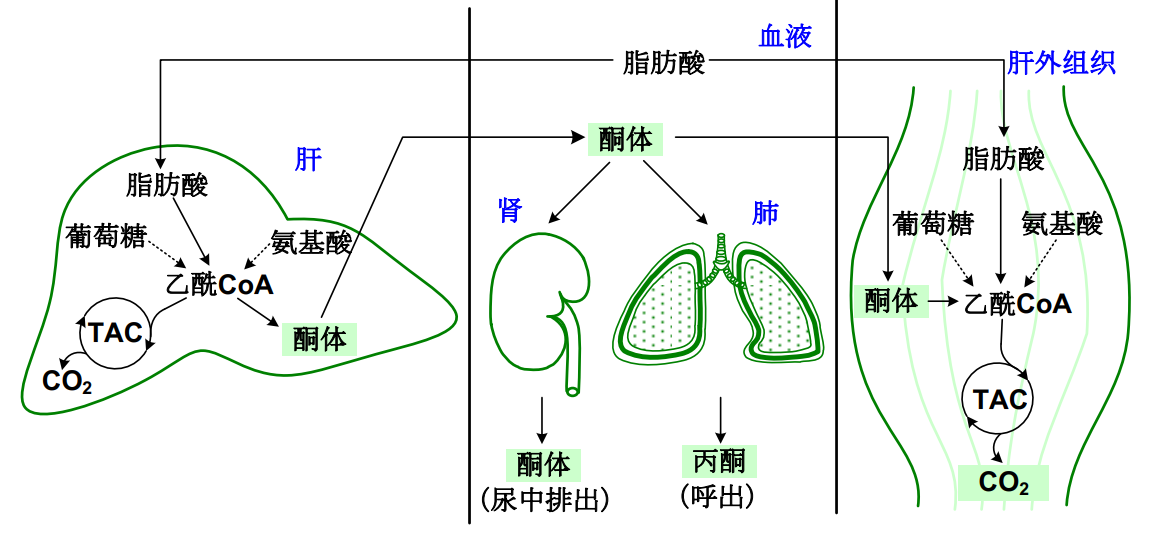
值得注意的是,虽然酮体在肝脏产生,酮体的利用在肝外进行。
酮体的正常血浆水平:0.03~0.5mmol/L (0.3~5mg/dL)。
酮体的生成途径和种类
酮体的利用
肝外组织虽然不能产生酮体,但是可以高效地利用酮体(存在相应活性的酶):
- 琥珀酰CoA转硫酶:主要存在于心、肾、脑和骨骼肌细胞的线粒体中。
- 乙酰乙酸硫激酶:主要存在心、脑和肾细胞线粒体中
将酮体转变为乙酰CoA进入到TCA循环中。
酮体生成的生理意义
酮体是肝脏输出能源物质的一种形式,在长期饥饿时是脑和肌肉的主要能源物质。
正常血酮体含量为0.03~0.5mmol/L,在饥饿、糖代谢障碍或供糖不足的情况下,肝脏产生大量酮体,使得血液中酮体含量升高,被称为高血酮症。
甘油三酯的合成代谢
甘油三酯的合成可以分为两大阶段:脂酸合成和甘油三酯的合成两个部分。
脂肪酸的合成碳源主要来自糖代谢与氨基酸代谢生成的乙酰CoA,主要在细胞液中进行。
饱和脂肪酸的合成代谢
饱和脂肪酸主要在肝脏、脂肪等组织的胞液和肝内质网中进行。前者主要合成16碳的软脂酸/棕榈酸,后者主要负责碳链延长。
合成原料:
- 乙酰CoA(主要来自葡萄糖)
- NADPH (主要来自磷酸戊糖途径)
- ATP、\(\ce{CO2}\)和\(\ce{Mn^{2+}}\)等等
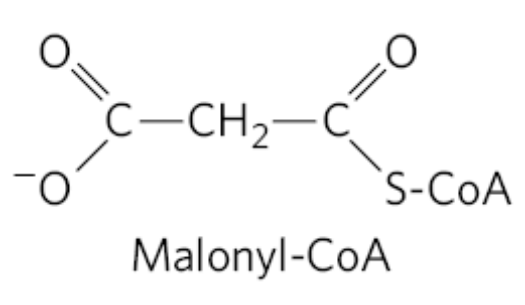
在合成之前需要先将乙酰CoA活化为丙二酸单酰CoA。
乙酰CoA转运途径
与丙酮酸进入线粒体类似,乙酰CoA通过线粒体内膜也需要特定转运体系——柠檬酸-丙酮酸循环。
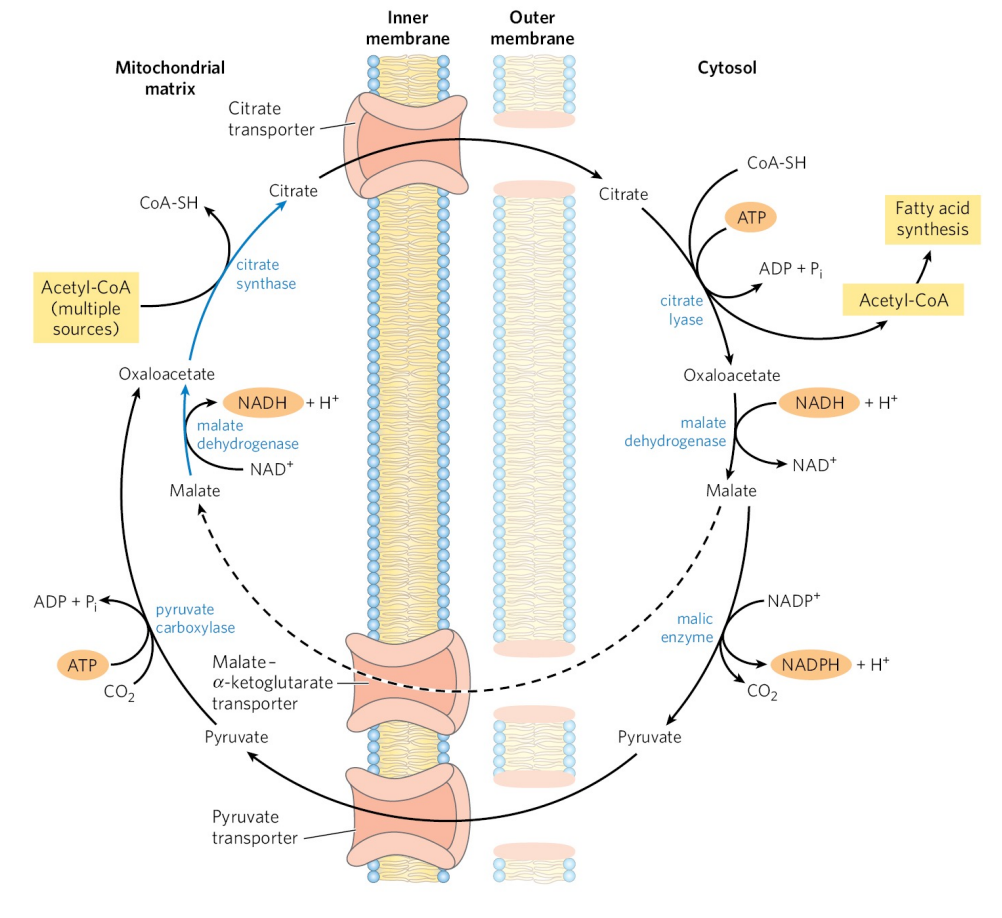
在上述穿梭体系中,苹果酸-丙酮酸-草酰乙酸的回归途径也为胞质提供了一分子的NADPH,和磷酸戊糖途径一起为之后的脂肪链合成准备原料。
活化阶段
在活化阶段,乙酰CoA被乙酰CoA羧化酶(ACC)催化生成丙二酸单酰CoA。
该酶存在于细胞质中,有无活性单体和有活性多聚体两种形式。
Note
所有羧化酶都需要生物素作为辅基。
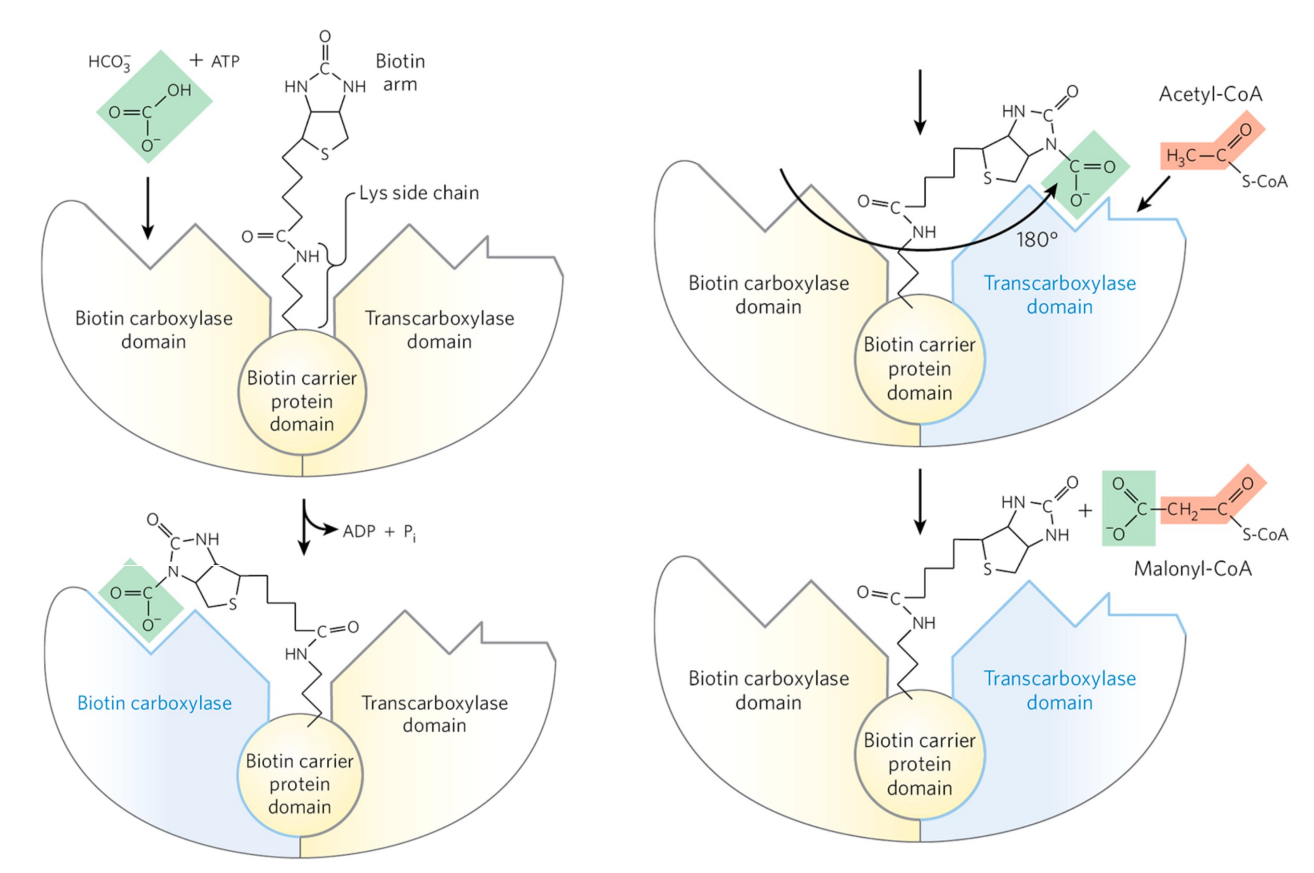
ACC在激活后会形成多聚体,它接受如下调节方式:
- 变构调节:
- 变构激活剂:柠檬酸与异柠檬酸
- 变构抑制剂:长链脂酰CoA
- 化学修饰条件:磷酸化抑制,去磷酸化激活
- 上游调节酶:AMPK激酶(胰高血糖素)磷酸化抑制ACC、蛋白磷酸酶(胰岛素)去磷酸化激活ACC
- 高AMP水平(能量匮乏)也会抑制ACC活性